Lipid signalling
Membrane functions:
barrier
signal transduction
transport
sets up gradients
cell shape and rigidity
compartmentalisation and specialisation
Membrane structure:
4nm, 2 molecules thick
lipids and proteins
mainly hydrophobic, lipids amphipathic
held together by weak non-covalent interactions (strong collectively)
fluid
asymmetric (outer and inner leaflet different in composition)
Hydrophobic effect: indirect effect resulting from the constant rapid exchanging of hydrogen bonds between water molecules. Where water interacts with a non H bonding group eg methyl, water molecules have fewer opportunities for H bond exchange so H bonds remain intact for longer creating a more solid state at the interface and decreased entropy as a result. Water at interface rotationally and translationally constrained. Minimising area of contact between water and non polar groups eg hydrocarbon regions of proteins increases entropy. This occurs through clustering non-polar groups together.
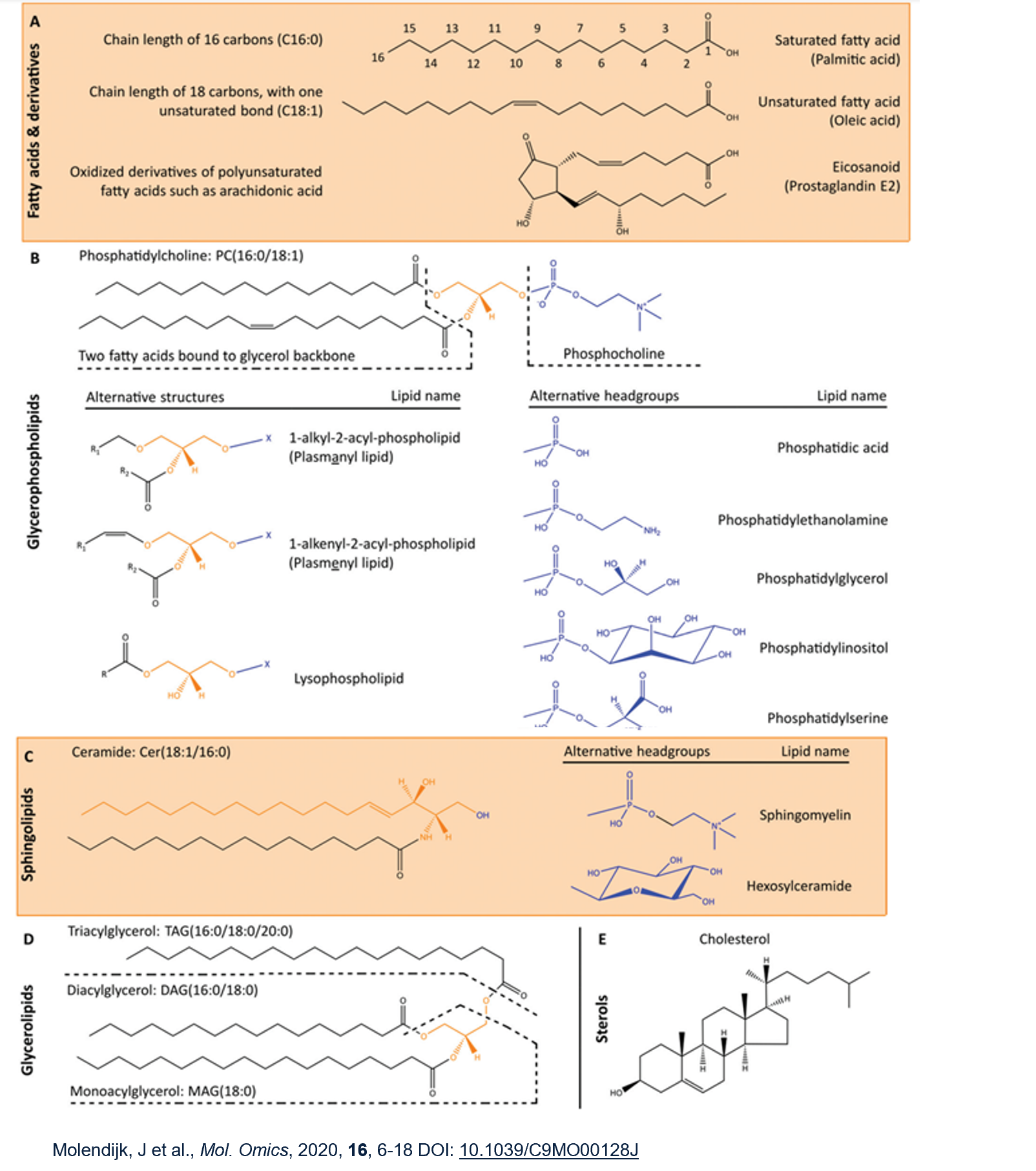
Lipids are a greatly diverse group divided into classes eg fatty acids, glycerophospholipids, sphingolipids, glycerolipids and sterols. Need to be able to recognise these classes.
Glycerophospholipids = 2 fatty acids linked to glycerol with phospho head group, most common being phosphatidylcholine
Sphingolipids have sphingoid base backbone instead of glycerol. Sphingoid = aliphatic amino alcohols. Associate with each other and with cholesterol/other proteins.
Glycerolipids have at least one FA chain but no headgroup. Intermediates in GP synthesis, lipid storage or signalling functions.
Sterols eg cholesterol have a more rigid structure.
The three types of membrane lipids are phospholipids, glycolipids and cholesterol. The majority are synthesised in the ER and are sometimes used as drug targets. They are all amphipathic.
Phospholipids are able to spontaneously form vesicles due to the hydrophobic effect.
Asymmetry in the bilayer:
Maintenance of an asymmetric bilayer is important for many different reasons eg in signalling, apoptosis, blood clot formation and protein targeting, sorting and function.
Signalling: Phosphatidylinositol is a key signalling molecule on the INSIDE of the membrane.
Apoptosis: during apoptosis a flippase specific to phosphatidylserine is activated revealing the lipid on the surface of the dying cell rather than on the inside. This means the dying cell is recognised by macrophages which consume it.
Blood clot formation: when platelets are activated by blood vessel injury, they scramble the lipids in the two membrane leaflets to expose phosphatidylserine (PS) at the surface where it acts as a cofactor in converting coagulation proteins to a blood clot. Normally, platelets sequester PS in their cytoplasmic leaflet.
Cell structure: there's asymmetry in lipid composition among the organelles.
Lipids are a major research target of the post-genomic revolution. They function as signalling molecules, chemical identifiers of specific intracellular compartments and as intracellular energy stores.
Phospholipases:
found everywhere, diverse in hydrolysis sites
hydrolyses phospholipids into fatty acids and other lipophilic substances
hydrolysing membrane lipids removes them from membrane, potentially changing membrane structure, integrity and fluidity
lipid derivative sometimes released, acts as signalling molecule eg cytosolic proteins
cleave glycerophospholipids → lipid 2nd messengers
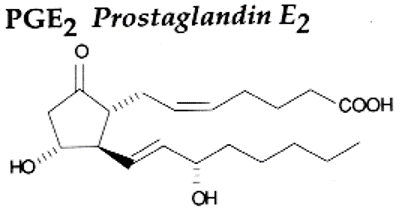
Phospholipase A2 has a major role in signal transduction as it releases arachidonic acid (C20H32O2) which is a precursor in biosynthesis for prostaglandins, thromboxanes and leukotrienes. It is of interest as a pharmaceutical target due to its role in inflammation mediators.
Prostaglandins:
paracrine (locally active) or autocrine (act on same cell it was synthesised by) function
potent
short half life before inactivation/excretion
9 known prostaglandin receptors on different cell types eg platelets, endothelium, uterine and mast cells
act on GPCRs
Arachidonic acid is converted to prostaglandins H2 by COX1 enzymes. Attached synthase enzymes convert prostaglandin H2 into TXA2, prostaglandins or prostacyclin.
Prostaglandin functions:
dilation in vascular smooth muscle cells
platelet aggregation
sensitise spinal neurons to pain
constrict smooth muscle (uterine contraction eg mifepristone which induces abortion)
regulate inflammatory processes
regulates calcium movement
Thromboxane is a vasoconstrictor and hypertensive agent which facilitates platelet aggregation. It’s in homeostatic balance with prostacyclin.
Leukotrienes stimulate proinflammatory activity eg endothelial cell adherance and production of chemokines by mast cells. They also induce asthma and other inflammatory disorders.
Endocannabinoid system:
important neuromodulatory system
comprises cannabinoid receptors, endogenous cannabinoids and enzymes which synthesise and degrade endocannabinoids
disruptions in ECS linked to schizophrenia and other conditions
Examples of endogenous cannabinoids are 2-arachidonoyl glycerol (2-AG) and arachidonoyl ethanolamids (anandamide, AEA). They both contain arachidonic acid and are synthesised from omega 6 polyunsaturated fatty acids but have very different routes of synthesis and degradation in vivo, mediated by different enzymes.
Cannabinoid receptors CB1 and CB2 are both GPCRs which couple to Gi and Go classes. CB1 receptors are found in CNS presynaptic neurones whereas CB2 is found in the peripheral nervous system and immune cells. Other receptors include transient receptor potential channels and peroxisome proliferator activated receptors.
ECS functions are complex. It is linked with appetite, metabolism, pain, inflammation, sleep, motor control and more. Rimonabant is an inverse agonist of CB1 receptors developed as an anti-obesity drug. It was very effective but had dramatic effects on mood so was removed on an emergency basis.
Phosphoinositides:
many phosphorylated forms
localised to distinct cellular components eg ptdins3P to endosomes, Ptdins4P to ER, Golgi and secretory vesicles, PtdIns(3,5)P2 to MVB/late endosome and PtdIns(4,5)P2 to membrane clathrin coated pits
minor component of total lipidome
signalling
intracellular trafficking
compartment/organelle identity
many important biological roles eg kinases and phosphatases
inter-related species involved in a cycle of non-vascular transfer of prostaglandins
Example phosphoinositides are phosphoinositide 3-kinases. PI3Ks are involved in many biological responses such as vesicle trafficking, glucose transport, chemotaxis and DNA synthesis. They can have many different effects in the same cell. This is due to spatial and temporal regulation, and different effector complements. Some cytosolic proteins have domains which bind polar lipid heads in the membrane eg Plechstrin Homology domains bind selectively to phosphorylated phosphatidylinositol derivatives eg PI3P. PI3K is able to modulate affinity of cytosolic proteins for the membrane surface. This means cells might accumulate specific molecules recruited to the cell surface upon PI3K activation.
Pi3P is generated in fat cells in response to insulin and PDGF. PDGF causes localised clusters of PI3P generation whereas insulin causes mass PI3P generation.
Interconnected nature of lipid signalling:
Lipids can influence membrane-protein interfaces in all cells and cell activity differs due to lipid/protein/enzyme composition. 50% are non polar eg cholesterol which is key for membrane stability and found highly concentrated in lipid rafts. Cholesterol interacts with proteins to support signalling eg GPCRs. The other 50% of lipids are polar such as glycerophospholipids and sphingolipids. Understanding lipid interconnectivity and behaviour adds to complexity of signalling cascades.
Lipid rafts:
Bilayer consists of raft and non-raft regions. The raft regions are thicker due to more elongated lipids and high sphingolipid and cholesterol content. Proteins with different transmembrane domain lengths may selectively populate different areas of the membrane. Cholesterol is enriched in the inner leaflet of rafts. Different proteins may be recruited by different mechanisms to collect specific signalling molecules in a particular area making the signalling process more efficient.