neuromuscular
The Motor Unit of Skeletal Muscle
Somatic Motor Neurons
All skeletal muscles are under voluntary or reflex control by motor neurons of the somatic motor system.
Somatic motor neurons are efferent neurons.
Their cell bodies are located in the central nervous system (CNS).
For spinal nerves, the cell body resides in the ventral horn of the spinal cord.
Pathway of a Motor Neuron
1. Cell Body
Located in the ventral horn of the spinal cord.
2. Axon Leaves the Spinal Cord
The motor neuron sends out an axon toward the skeletal muscle.
3. Axon Branches
Near its termination, the axon typically branches.
These branches innervate a few or many individual muscle fibers.
A single muscle fiber responds to only one motor neuron.
Motor Unit
Definition
A motor unit =
one motor neuron
all the muscle fibers innervated by its branches
So:
one neuron → many muscle fibers
all of them together = one motor unit.
Motor Neuron Pool
A muscle does not receive input from only one motor neuron.
A typical skeletal muscle receives innervation from ~100 somatic motor neurons.
The collection of many motor neurons supplying one muscle is called a motor neuron unit.
The Innervation Ratio of the Skeletal Muscle
Whole Muscle Function
A whole muscle can produce:
a wide range of forces
a graded range of shortening
by varying the number of motor units excited within the muscle.
Innervation Ratio
Definition
The innervation ratio of a whole skeletal muscle is defined as:
the number of muscle fibers innervated by a single motor neuron
Small Innervation Ratio
Function
Muscles with a small innervation ratio control:
fine movements
small forces
Example
Fine, high precision movements of the extraocular muscles that control positioning movements of the eye are achieved through an innervation ratio of:
~3 muscle fibers per neuron
Large Innervation Ratio
Function
Muscles with a large innervation ratio control:
rough movement
development of large forces
Examples
Soleus Muscle
Postural control by the soleus muscle uses an innervation ratio of:
~200
Gastrocnemius Muscle
The gastrocnemius muscle, which is capable of developing large forces required in athletic activities such as jumping, has innervation ratios that vary from:
~100 to ~1000
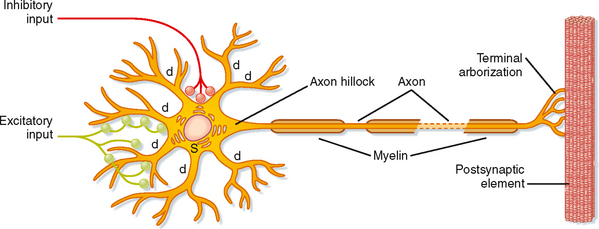
Neuromuscular Junction / Motor End-Plate
Motor Neuron Pathway
motor neuron (spinal cord)
→ one axon
→ branches into axon processes near the point of contact with the target muscle
Axon Branch
one axon branch innervate an individual muscular fiber
through:
Terminal Arborizations
a small tree-like patch of unmyelinated nerve processes
Nerve Endings
ending into bulb-shaped terminals called boutons
Schwann Cells
Schwann cells intimately associate with the nerve terminal
form a cap over the face of the nerve membrane
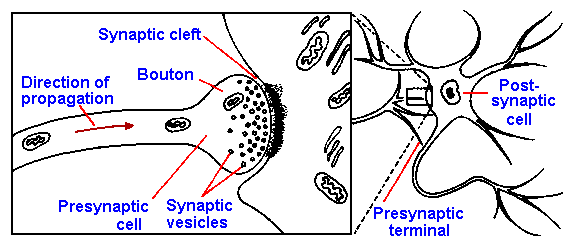
Synaptic Transmission at the Neuromuscular Junction – Motor End-Plate
Presynaptic Boutons
presynaptic boutons contain ACh vesicles
6000–10000 ACh molecules / vesicle = quantum
→ quantum release into the synaptic cleft
After Each AP
within a few sec. after each AP, “coated pits” appear in the presynaptic membrane
caused by clathrin = contractile proteins in the nerve endings
within ~20 sec. the proteins contract and cause the pits to break away to the interior of the membrane
thus forming new vesicles
Vesicle Reloading
bouton reloads its discharged synaptic vesicles by resynthesizing ACh
transport it into the vesicles through an ACh-H+ exchanger
working on a vacuolar H+-pump expense
AChE
AChE = acetylcholinesterase, in the synaptic cleft
Breakdown of ACh
AChE + ACh
↓
acetyl-AChE
↓
acetate + AChE
Products
choline
acetate
Neuromuscular Junction
Synaptic Cleft
synaptic cleft is ~50 nm wide
filled with a meshwork of proteins and proteoglycans
part of the extracellular matrix
Muscle Basement Membrane
Synaptic Basal Lamina
Contains Proteins
collagen
laminin
Functions
mediate adhesion of the neuromuscular junction
play important roles in:
synapse development
regeneration
Acetylcholinesterase (AChE)
synaptic basal lamina contains a high concentration of acetylcholinesterase (AChE)
terminates synaptic transmission
rapidly hydrolyzing free ACh to:
choline
acetate
Postsynaptic Membrane
postsynaptic membrane of the skeletal muscle fiber (motor end-plate)
lying directly under the nerve terminal
characterized by extensive invaginations/infoldings
= post-junctional folds
Function
increase postsynaptic surface
Postsynaptic Receptors
postsynaptic nicotinic ACh receptors
ionotropic receptors
→ end-plate potential (EPP)
= excitatory postsynaptic potential (EPSP)
Neuromuscular Junction
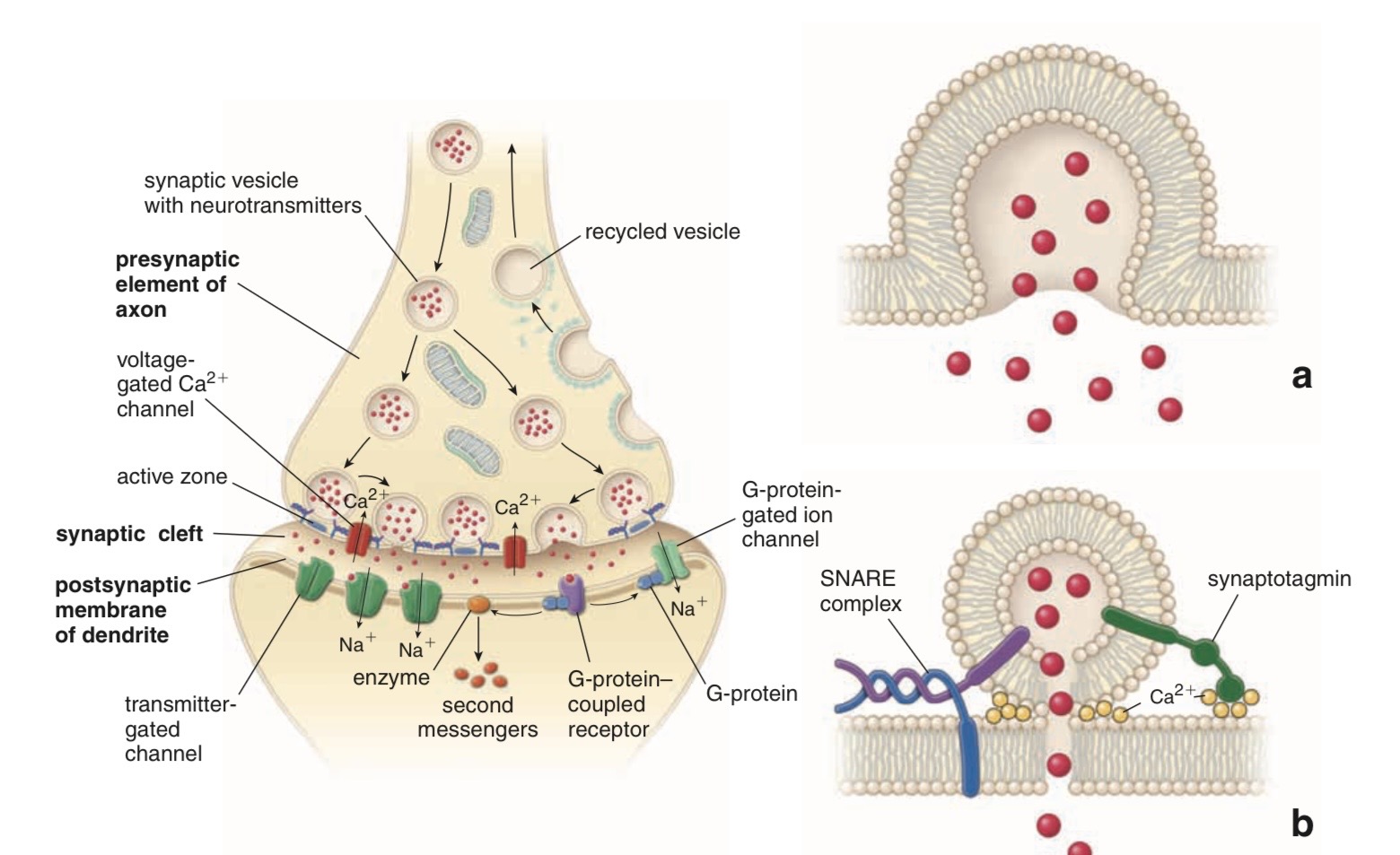
Action Potential (AP)
AP spreads over the presynaptic terminal
↓
voltage-gated Ca channels open
↓
Ca2+ influx
↓
ACh vesicles draw to the presynaptic membrane
↓
vesicles fuse with the presynaptic membrane
↓
ACh emptied into the synaptic space by exocytosis
Removal of ACh
ACh is removed rapidly (few msec) by:
1. Acetylcholinesterase
attached mainly to the spongy layer of fine connective tissue in the synaptic space
2. Diffusion
diffusion out of the synaptic space
a smaller amount
ACh Nicotinic Receptors
ACh nicotinic receptors in the muscle fiber membrane are ion channels
diameter ~0.65 nm
Allow
positive ions:
Na+
K+
to move easily
Cl-
repelled by the negative charge of the channel pore
End-Plate Potential (EPP)
Na+ influx creates a local positive potential change of 50–75 mV inside the muscle fiber membrane
= end-plate potential (EPP)
↓
initiates a regenerating AP
↓
AP spreads along the muscle membrane
↓
muscle contraction
Synaptic Facilitation
short-lived enhancement of the EPP in response to:
a brief increase in the frequency of nerve stimulation
a transient increase in the mean number of quanta/nerve stimulus
Synaptic Potentiation
(or post-tetanic potentiation)
long-lived and pronounced increase in ACh release
occurs after a long period of high-frequency nerve stimulation
can last for minutes after the conditioning stimulus
may be caused by a period of intense nerve firing
increases [Ca2+]i in the presynaptic terminal
thus increases the probability of exocytosis
Synaptic Depression
transient decrease in the efficiency of transmitter release
→ reduction in the EPP in response to a period of frequent nerve stimulationmay result from temporary depletion of transmitter-loaded vesicles from the presynaptic terminal
→ reduction in the number of available quanta.
Pharmacology of the Vertebrate Neuromuscular Junction (NMJ)
Presynaptic
Neuronal Na+ Channel
Prevent Depolarization
Tetrodotoxin
Saxitoxin
Ca2+ Channel
ω-Conotoxin
K+ Channel
Inhibit Repolarization
Dendrotoxin
ACh Release
Tetanus toxin
Botulinum toxin
Postsynaptic
Muscle Na+ Channel
Tetrodotoxin
Saxitoxin
μ-Conotoxin
AChR Channel
Agonists (+)
Acetylcholine
Nicotine
Antagonists (−)
d-Tubocurarine
α-Bungarotoxin
Acetylcholinesterase
Physostigmine
DFP
Notes
many proteins involved in synaptic transmission at the mammalian neuromuscular junction are targets of naturally occurring or synthetic drugs
antagonists are shown as minus signs highlighted in red
agonists are shown as plus signs highlighted in green
Drugs that Enhance or Block Transmission at the Neuromuscular Junction
Drugs that Stimulate the Muscle Fiber by Acetylcholine-Like Action
Drugs
methacholine
carbachol
nicotine
Characteristics
ACh agonists
not destroyed by cholinesterase
ordestroyed so slowly that their action persists for many minutes to several hours
Effects
cause localized areas of depolarization of motor end plate
every time the muscle fiber recovers from a previous contraction, these depolarized areas, by virtue of leaking ions, initiate a new AP
→ muscle spasm
Drugs that Stimulate the Neuromuscular Junction by Inactivating Acetylcholinesterase
Drugs
neostigmine
physostigmine
pyridostigmine
diisopropyl fluorophosphate
Mechanism
inactivate AChE
with each successive AP, additional ACh accumulates
stimulates the muscle fiber repetitively
→ muscle spasm caused by minimum stimulation
can also cause death due to laryngeal spasm
Neostigmine and Physostigmine
combine with AChE
reversibly inactivate the AChE
for up to several hours
Pyridostigmine
used in myasthenia gravis treatment
Diisopropyl Fluorophosphate
powerful “nerve” gas poison
inactivates AChE for weeks
particularly lethal poison
Drugs that Inhibit the Postsynaptic Transmission at the Neuromuscular Junction
Curariform Drugs
prevent passage of impulses from the nerve ending into the muscle
by competing for the ACh receptor sites
Example
D-tubocurarine
blocks the action of ACh on the muscle fiber ACh receptors
prevents sufficient increase in permeability of the muscle membrane channels
prevents initiation of an action potential
Myasthenia Gravis
= muscle weakness
(from the Greek mys and asthenia)
Disorder
acquired autoimmune disorder
spontaneous production of anti-ACh receptor (AChR) antibodies
Effects
progressive loss of muscle AChRs
degeneration of post-junctional folds
Antibody Target
most common target:
region of the AChR α subunit
called MIR (main immunogenic region)
Clinical
fatigue
weakness of skeletal muscle
Severe Cases
paralysis of the respiratory muscles
→ death
Treatment
1. Reduce the Potency of the Immunological Attack
Immuno-suppressants
corticosteroids
Plasmapheresis
removal of antibodies from the patient’s serum
2. Enhance Cholinergic Activity Within the Synapse
AChE Inhibitors
pyridostigmine
Important
dosage must be carefully monitored
to prevent overexposure of the remaining AChRs to ACh
Overstimulation Effects
overstimulation of the postsynaptic receptors
prolonged depolarization of the postsynaptic membrane
inactivation of neighboring Na+ channels
synaptic blockade
3. Thymoma Removal
some patients with myasthenia gravis have a thymus gland tumor
removal of the thymoma leads to clinical improvement in nearly 75% of the cases
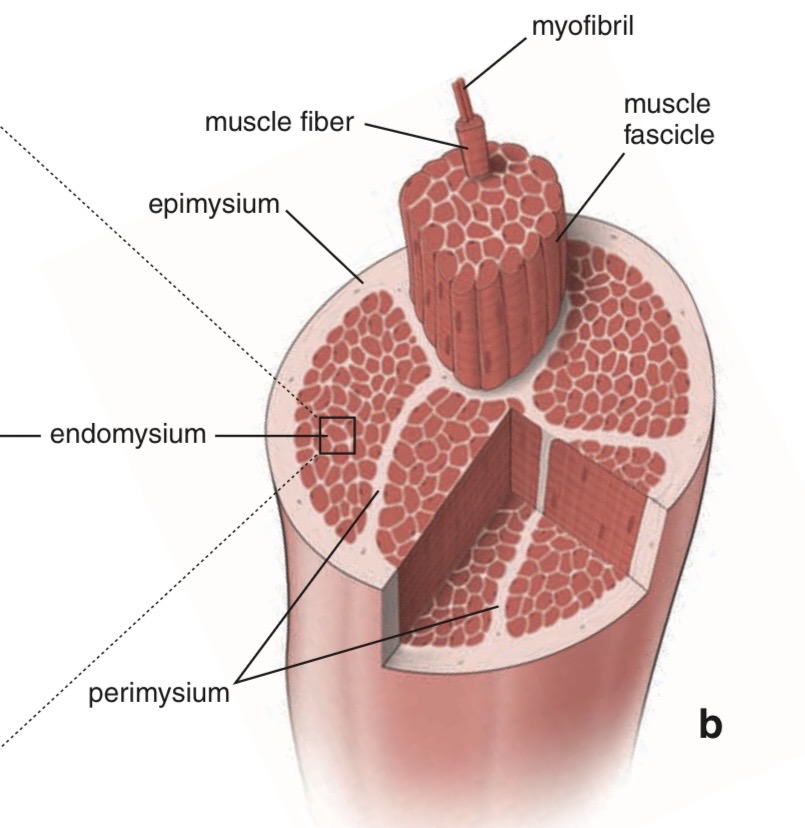
Muscle Fiber Cell / Myofiber
smallest contractile unit of skeletal muscle
multinucleated
elongated cell
Endomysium
single muscle fibers are surrounded by a sheath called the endomysium
Sarcolemma
beneath the endomysium surrounding each muscle fiber is sarcolemma
plasma membrane of the muscle cell
Fascicle
a bundle of linearly aligned muscle fibers forms a fascicle
enveloped by a sheath called the perimysium
Muscle
bundles of fascicles form a muscle
covered by epimysium
extends towards tendons
Myofibrils
an individual skeletal muscle cell contains a densely arranged parallel array of cylindrical elements called myofibrils
Sarcomeres
myofibrils
= an end-to-end chain of regular repeating units
= sarcomeres
Myofilaments
sarcomeres consist of smaller interdigitating filaments
= myofilaments
Types
actin
myosin
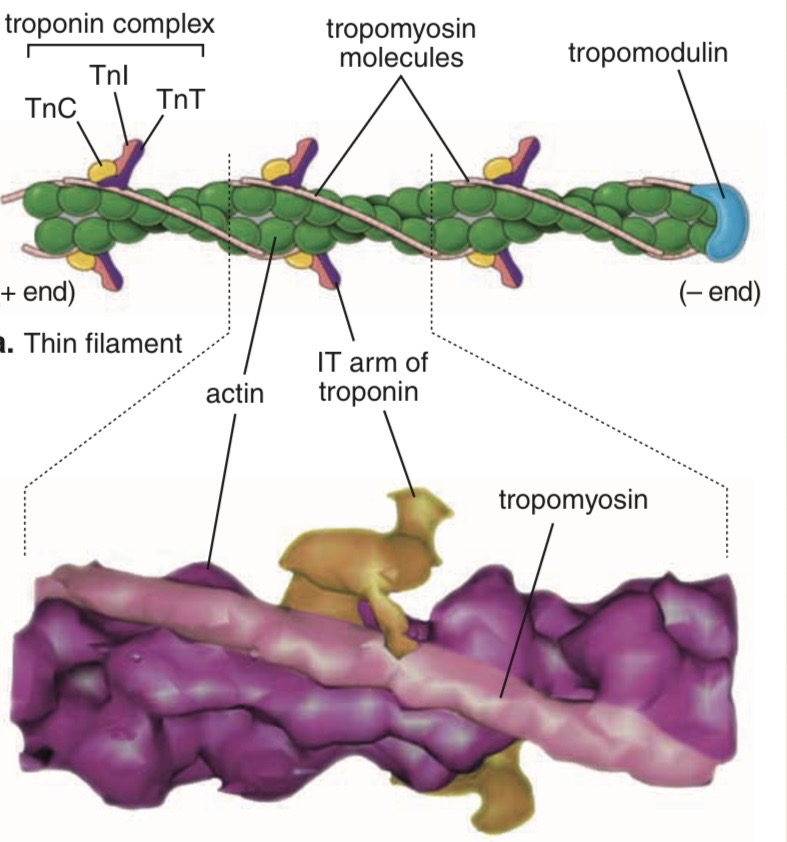
Tropomyosin ( thin filament )
two identical α-helices
coil around each other
sit near the two grooves formed by the two helical actin strands
Head-to-Tail Contact
head-to-tail contact between neighboring tropomyosin molecules
results in two nearly continuous helical filaments
shadow the actin double helix
Role of Tropomyosin
regulate the binding of myosin head groups to actin
Troponin
heterotrimer consisting of:
1. Troponin T (TnT or TNNT)
binds to a single molecule of tropomyosin
2. Troponin C (TnC or TNNC)
binds Ca2+
related to another Ca2+-binding protein:
calmodulin
3. Troponin I (TnI or TNNI)
binds to actin
inhibits contraction
Coordinated Interaction
coordinated interaction among:
troponin
tropomyosin
actin
allows:
actin-myosin interactions to be regulated by changes in [Ca2+]i
Thick Filaments
10 nm in diameter
1.6 micrometers in length
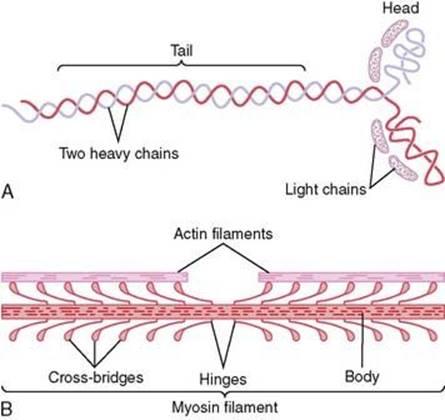
Myosin
bipolar assembly of multiple myosin II molecules
Structure of Myosin
Heavy Chains
2 intertwined heavy chains
contain:
tail
hinge
head region
Light Chains
2 alkali (or essential) light chains
2 regulatory light chains
Heads of the Heavy Chains
2 globular heads of the heavy chains
= crossbridges between the thick and thin filaments of the sarcomere
Binding Sites
Actin Binding Site
site for binding actin
ATP Binding Site
site for binding and hydrolyzing ATP
Light Chain Complex
forms a complex with two light chains:
one regulatory
one alkali
Regulatory Light Chain
regulates the ATPase activity of myosin
Alkali Light Chain
stabilizes the myosin head region
Regulation of Myosin Regulatory Light Chain
regulated through phosphorylation
by:
Ca2+-dependent kinases
Ca2+-independent kinases
Calcium Switch for Skeletal Muscle Contraction
When Ca²⁺ is LOW (< 10⁻⁹ M)
No Ca²⁺ binds to troponin
Tropomyosin blocks the binding sites on actin
Myosin cannot interact with actin
Contact between binding sites is inhibited
Muscle is relaxed
When Ca²⁺ is HIGH (> 10⁻⁵ M)
Ca²⁺ binds to troponin
Tropomyosin moves away from the binding sites
Myosin can interact with actin
Contact between binding sites is permitted
Muscle is contracted
Main Idea
Ca²⁺ ions control skeletal muscle contraction through the troponin–tropomyosin complex by regulating whether myosin heads can bind to active sites on actin.
Muscle Fiber Structure
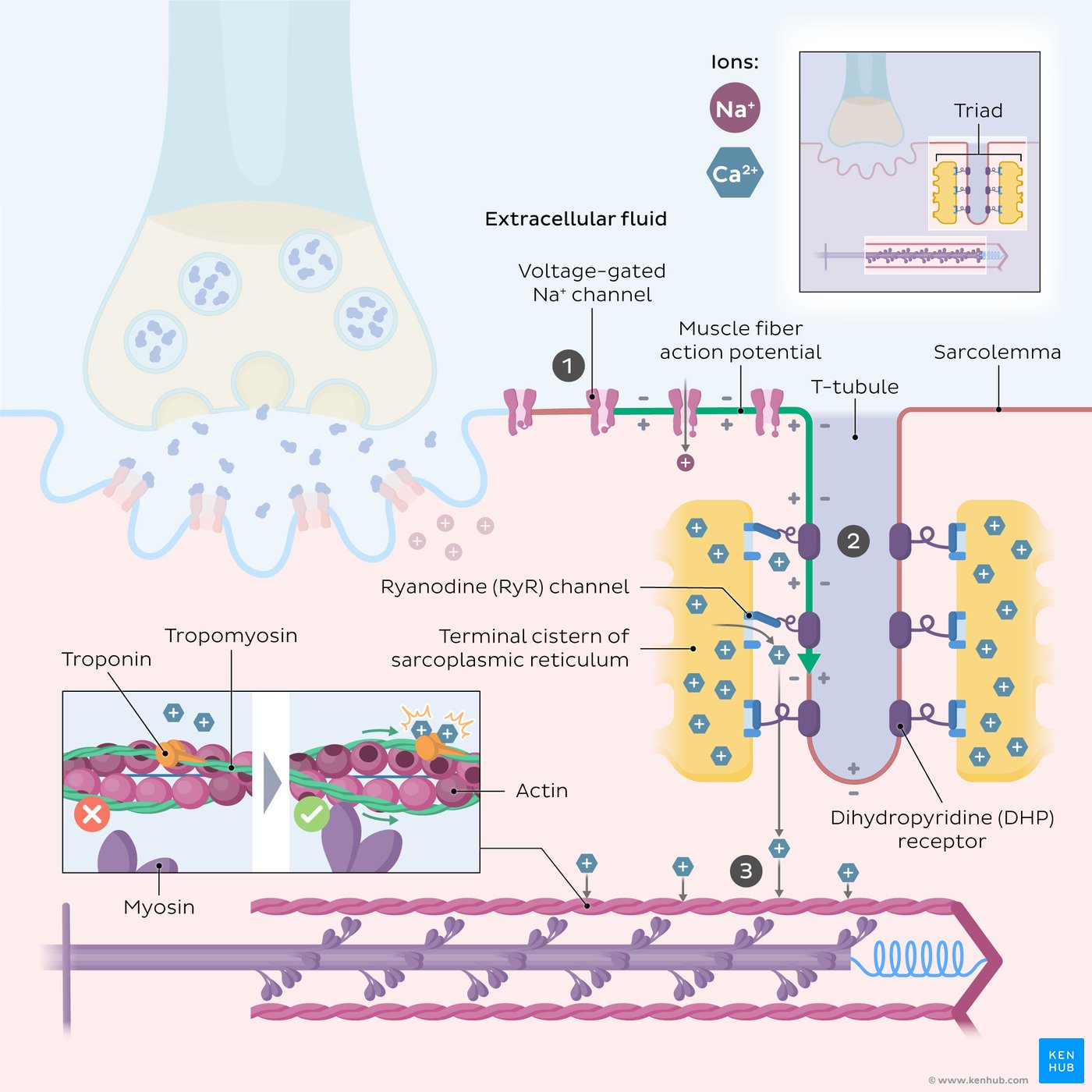
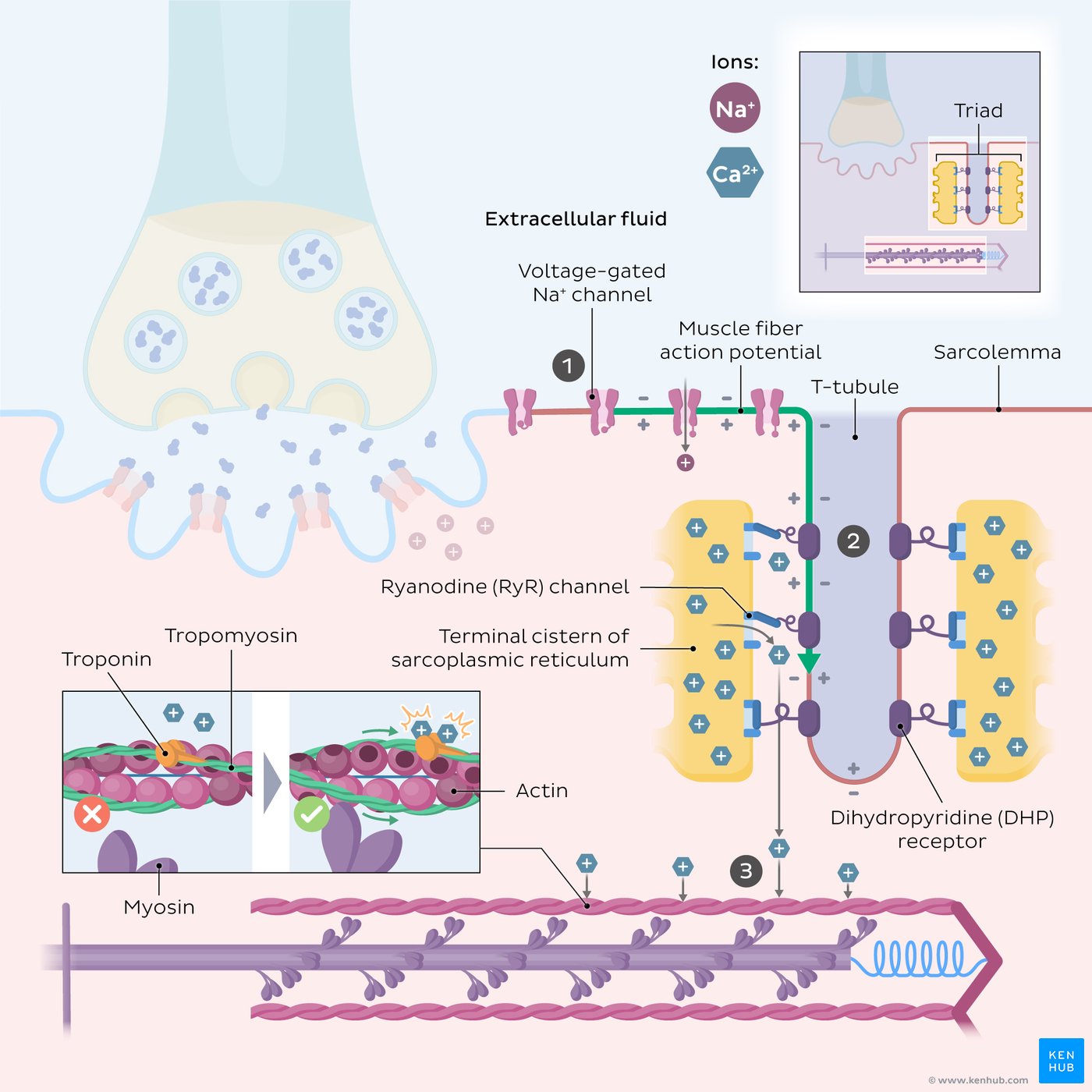
T Tubules
T tubules are tubular invaginations of the plasma membrane
Radially oriented
Plunge into the muscle fiber
Surround the myofibrils at the junctions of the A and I bands
Two T tubules in each sarcomere
Sarcoplasmic Reticulum (SR)
T tubules associate with two cisternae
Cisternae are specialized regions of the sarcoplasmic reticulum (SR)
SR serves as a storage organelle for intracellular Ca²⁺
Triad
A TRIAD consists of:
One transverse tubule (T tubule)
Two neighboring sarcoplasmic reticulum cisternae
Function
Important in coupling excitation to contraction in skeletal and cardiac muscle
Ca²⁺ Movement
Action potential travels along the cell membrane and T tubule
Causes Ca²⁺ release
Ca²⁺ translocation occurs
Ca²⁺ is later taken back up into the SR (Ca²⁺ reuptake).
Electrical Activity at the Neuromuscular Junction
Key Concept
Neuromuscular transmission occurs when a motor neuron action potential (AP) releases acetylcholine (ACh) at the neuromuscular junction, producing depolarization of the muscle end plate.
(A) Small Amount of Acetylcholine Released
Sequence
A motor axon AP reaches the axon terminal.
Small amount of ACh is released into the neuromuscular junction.
A small end plate depolarization occurs.
Depolarization spreads through the myoplasm by electrotonic conduction.
Electrotonic conduction occurs with decrement (signal weakens with distance).
Depolarization reaching the first voltage-gated Na⁺ channel is below threshold.
Na⁺ channels do not open.
No muscle action potential is formed.
Important Physiology Points (Exam Focus)
ACh causes inward membrane current.
End plate potential spreads electrotonically.
Electrotonic conduction is:
passive
decremental
If depolarization is below Na⁺ channel threshold:
no AP
no muscle fiber excitation
(B) Greater Frequency of Motor Axon APs
Sequence
Many motor axon APs arrive at the terminal.
More ACh is released.
Higher ACh concentration in the neuromuscular junction.
Larger end plate potential (EPP) is produced.
Depolarization still spreads with decrement.
Depolarization reaching first Na⁺ channels is now above threshold.
Voltage-gated Na⁺ channels open.
Muscle fiber generates a muscle AP.
Muscle AP is:
self-reinforcing
propagated without decrement
AP travels along the muscle fiber similar to conduction in unmyelinated nerve fibers.
Important Physiology Points (Exam Focus)
Increased AP frequency → increased ACh release.
Larger EPP increases likelihood of reaching threshold.
Threshold opening of Na⁺ channels is essential for muscle AP generation.
Muscle AP propagation is:
regenerative
non-decremental
High-Yield Final Exam Notes
Definitions
End Plate Potential (EPP)
Local depolarization at the motor end plate caused by ACh.
Electrotonic Conduction
Passive spread of depolarization with gradual decrease in amplitude.
Threshold
Minimum depolarization needed to open voltage-gated Na⁺ channels.
Depolarization of the T-Tubule Membrane Results in Ca²⁺ Release from the Sarcoplasmic Reticulum
Main Concept
Action potential (AP) propagates into the T tubules → activates voltage-gated L-type Ca²⁺ channels (DHP receptors) → activates SR Ca²⁺-release channels (RYR1) → Ca²⁺ released from sarcoplasmic reticulum (SR).
Step-by-Step Mechanism
1. AP Propagation into T Tubules
Muscle action potential travels along sarcolemma.
AP enters T tubules.
T-tubule membrane becomes depolarized.
2. Triad Region
TRIAD =
1 transverse (T) tubule
2 terminal cisternae of sarcoplasmic reticulum
3. Activation of L-Type Ca²⁺ Channels
L-type Ca²⁺ Channels
Also called:
DHP receptors
Voltage-gated L-type Ca²⁺ channels
Located in:
T-tubule membrane
Function:
Activated by depolarization.
4. Conformational Change of DHP Receptor
Depolarization causes conformational change in DHP receptor.
This:
allows small Ca²⁺ entry into cell
mechanically activates SR Ca²⁺-release channels
5. Activation of Ryanodine Receptor (RYR1)
RYR1
Ca²⁺-release channel in SR membrane
Also called ryanodine receptor
Mechanism:
Activated by mechanical coupling with DHP receptor
Not dependent on extracellular Ca²⁺ entry
6. Result
Large amount of Ca²⁺ released from SR into cytoplasm.
Increased intracellular Ca²⁺ initiates muscle contraction.
Tetrad
Definition
Four voltage-activated L-type Ca²⁺ channels grouped together near one SR Ca²⁺-release channel.
This arrangement is called:
TETRAD
High-Yield Final Exam Notes
Must Memorize
AP in T tubule triggers Ca²⁺ release from SR.
DHP receptor = L-type voltage-gated Ca²⁺ channel.
RYR1 = SR Ca²⁺-release channel.
DHP receptor and RYR1 are mechanically coupled.
Skeletal muscle Ca²⁺ release does NOT depend on extracellular Ca²⁺ entry.
TRIAD:
1 T tubule
2 terminal cisternae of SR
Four DHP receptors form a tetrad.
Drug Associations
DHP Receptor
Inhibited by:
Dihydropyridines
Uses:
Ca²⁺ channel blockers
Antihypertensive drugs
Antiarrhythmic drugs
RYR1
Inhibited by ryanodine
Activated by caffeine
Flow Chart
AP in T tubule
→ DHP receptor activation
→ conformational change
→ RYR1 activation
→ Ca²⁺ release from SR
→ increased cytosolic Ca²⁺
→ muscle contraction
EC Coupling
Electrical excitation increases intracellular Ca²⁺.
Increased intracellular Ca²⁺ triggers contraction.
Sources of Ca²⁺
Extracellular space via voltage-gated Ca²⁺ channels
Sarcoplasmic reticulum (SR)
Excitation–Contraction (EC) Coupling in Skeletal Muscle
Main Concept
In skeletal muscle, EC coupling can occur without extracellular Ca²⁺.
Reason:
Direct mechanical coupling between:
L-type Ca²⁺ channel (DHP receptor)
SR Ca²⁺-release channel (RYR1)
This mechanism is called:
Electromechanical coupling
Sequence of EC Coupling
1. Motor Neuron Stimulation
Acetylcholine (ACh) released from motor neuron.
ACh binds receptors on motor end plate.
End plate potential generates muscle AP.
2. AP Propagation
Muscle AP spreads along sarcolemma.
AP enters T tubules.
3. DHP Receptor Activation
T-tubule depolarization activates L-type Ca²⁺ channels (DHP receptors).
4. Mechanical Activation of RYR1
DHP receptor mechanically activates RYR1 in SR membrane.
5. Ca²⁺ Release from SR
Ca²⁺ rapidly released from sarcoplasmic reticulum.
Rapid increase in intracellular Ca²⁺ concentration ([Ca²⁺]i).
6. Troponin C Activation
Ca²⁺ binds troponin C.
Tropomyosin moves away from myosin-binding sites.
7. Cross-Bridge Formation
Actin and myosin form cross-bridges.
Cross-bridge cycle begins.
Muscle contraction occurs.
8. Relaxation
Ca²⁺ actively transported back into SR.
Tropomyosin again blocks myosin-binding sites.
Muscle relaxes.
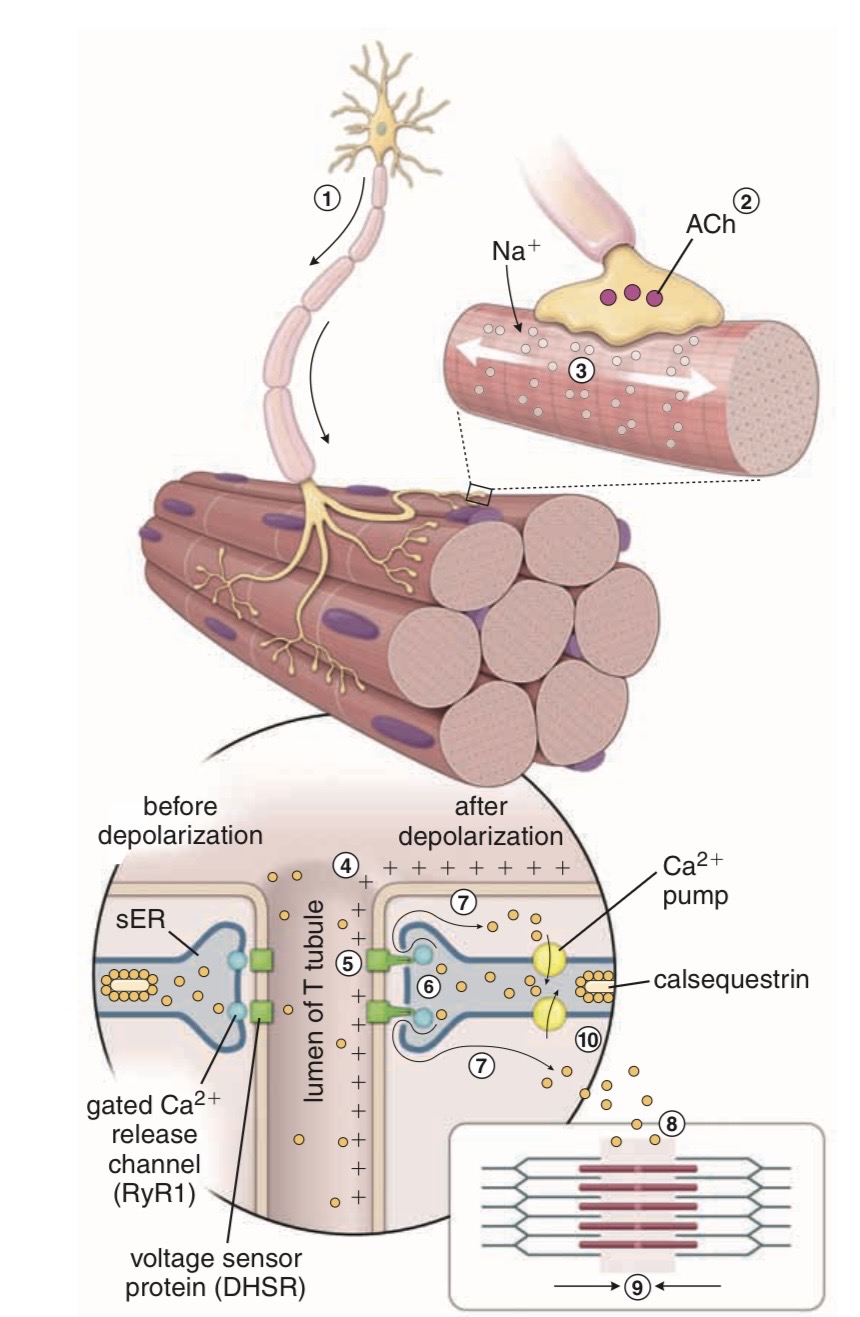
1. Motor Nerve Stimulates Muscle
A motor neuron sends an action potential (AP) to the muscle fiber.
At the nerve ending:
acetylcholine (ACh) is released.
ACh binds to:
nicotinic ACh receptors
on the muscle membrane.
2. Na⁺ Enters Muscle Fiber
When nicotinic receptors open:
large amounts of Na⁺ enter the muscle fiber.
This causes:
depolarization
If depolarization reaches threshold:
muscle action potential forms.
3. AP Spreads Through Muscle
The muscle AP spreads:
across the sarcolemma
down the T tubules
This carries the electrical signal deep into the muscle fiber.
4. EC Coupling Begins
Depolarization in T tubules activates:
DHP receptors (L-type Ca²⁺ channels)
DHP receptors are mechanically connected to:
RYR1 Ca²⁺-release channels
in the sarcoplasmic reticulum (SR).
This mechanical coupling opens RYR1.
5. Ca²⁺ Released from SR
The SR releases large amounts of stored Ca²⁺ into the cytoplasm.
Inside SR, Ca²⁺ is stored bound to:
calsequestrin
calreticulin
Now intracellular Ca²⁺ concentration rises rapidly.
This increased Ca²⁺ is the key signal for contraction.
6. Ca²⁺ Initiates Contraction
Ca²⁺ binds:
troponin C
This moves:
tropomyosin
away from actin binding sites.
Now:
myosin heads bind actin
Cross-bridges form between:
actin filaments
myosin filaments
7. Sliding Filament Mechanism
Myosin pulls actin filaments:
sliding alongside each other
This shortens the muscle fiber:
contraction
8. ATP Is Required
ATP is needed for:
cross-bridge cycling
muscle contraction
Ca²⁺ pumping back into SR
Without ATP:
contraction cannot continue
relaxation cannot occur
9. Relaxation
After contraction:
SERCA pump
actively pumps Ca²⁺ back into SR.
As Ca²⁺ leaves cytoplasm:
troponin loses Ca²⁺
tropomyosin blocks binding sites again
cross-bridges stop
Result:
muscle relaxation
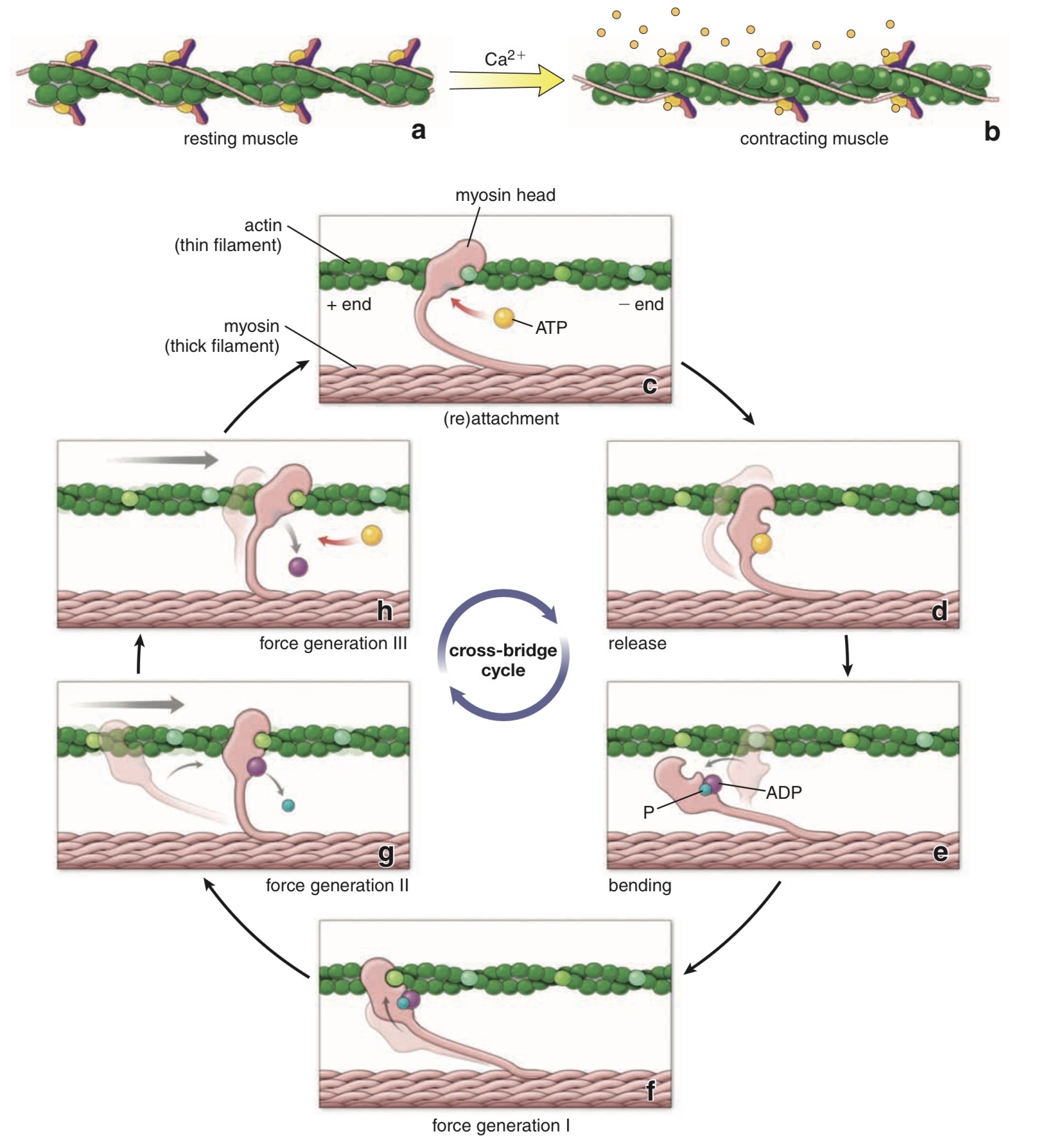
Cross-Bridge Cycle (Mechanism of Muscle Contraction)
This slide explains:
how myosin and actin interact to produce contraction.
The cycle repeats many times during contraction.
Step 1 — ATP Binding
ATP binds to:
myosin head
Effect:
decreases affinity of myosin for actin
myosin detaches from actin
Result:
cross-bridge breaks
If all myosin heads were detached:
muscle would be relaxed
Step 2 — ATP Hydrolysis
ATP is broken down:
ATP → ADP + Pi
This occurs on:
myosin head
The energy released causes:
myosin head to cock
Meaning:
myosin head pivots into high-energy position
about 90° angle
Important:
ADP and Pi remain attached to myosin head.
The cocked myosin head moves:
~11 nm along actin filament
Now it is ready to bind a new actin site.
Step 3 — Cross-Bridge Formation
Cocked myosin head binds:
actin filament
This forms:
actin–myosin cross-bridge
At this stage:
myosin still contains ADP + Pi
Step 4 — Power Stroke
Pi is released from myosin head.
This triggers:
power stroke
During power stroke:
myosin head bends about 45°
pulls actin filament toward center
Result:
force and movement generated
This is the main force-producing step.
Step 5 — ADP Release
After power stroke:
ADP leaves myosin head.
Now myosin remains tightly attached to actin:
rigid state
The cycle stops here until:
another ATP binds
New ATP binding:
detaches myosin
starts another cycle
Most Important Concept
ATP has TWO major roles:
1. Detaches myosin from actin
2. Cocks/energizes myosin head
Very High-Yield Exam Point
No ATP →
myosin cannot detach from actin
Result:
rigor state (rigor mortis)
Mechanism of Calcium Removal — Muscle Relaxation
This slide explains:
how Ca²⁺ is removed from cytoplasm to stop contraction.
Remember:
Muscle contraction requires ↑ intracellular Ca²⁺.
Relaxation requires ↓ intracellular Ca²⁺.
So after contraction, Ca²⁺ must be removed from cytoplasm.
Main Mechanisms Removing Ca²⁺
There are:
1. Sarcolemma mechanisms
(remove Ca²⁺ out of the cell)
2. SR mechanisms
(move Ca²⁺ back into SR)
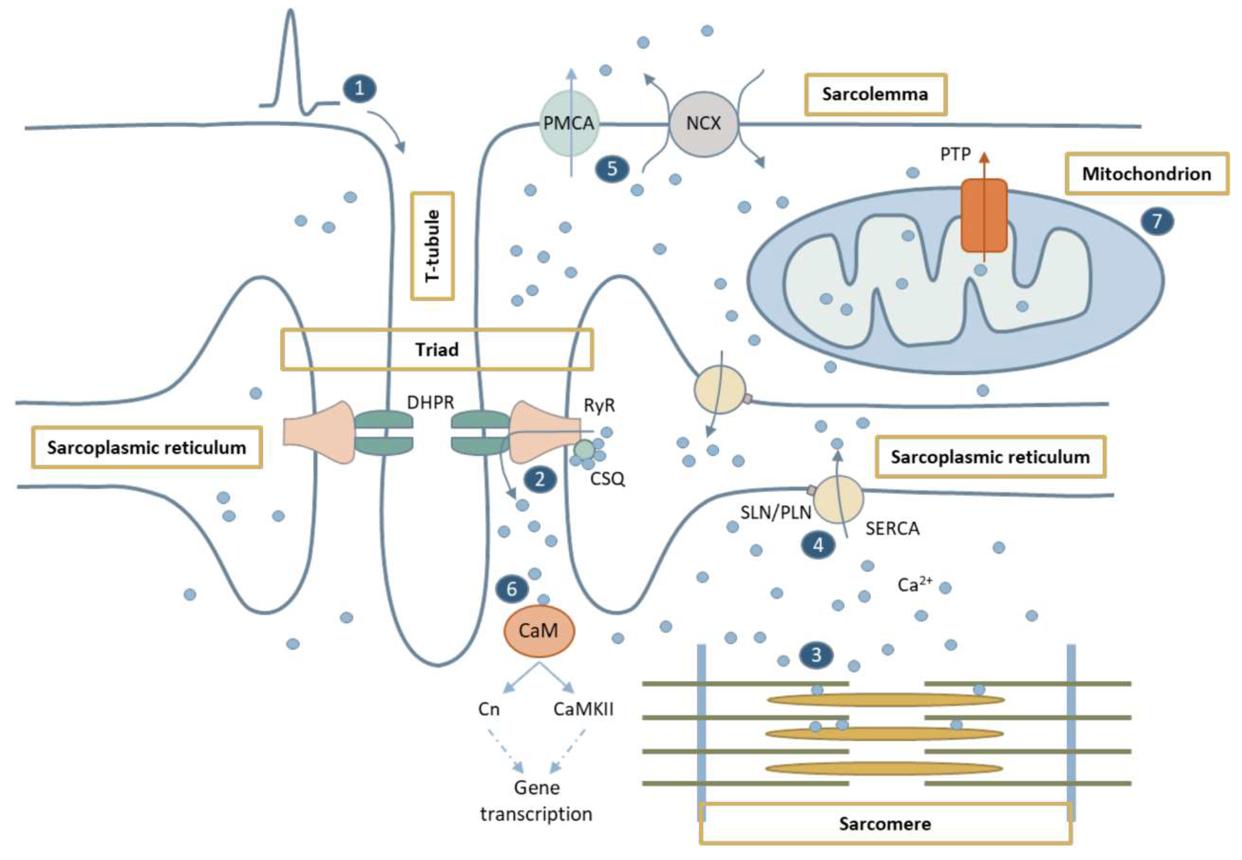
1. Sarcolemma Mechanisms
A. Na⁺–Ca²⁺ Exchanger (NCX)
Function:
removes Ca²⁺ from cell
exchanges intracellular Ca²⁺ for extracellular Na⁺
So:
Ca²⁺ goes out, Na⁺ comes in
B. Plasma Membrane Ca²⁺ ATPase (PMCA)
Function:
pumps Ca²⁺ out of cell
Important:
uses ATP
2. SR Mechanism — MOST IMPORTANT
SERCA Pump
Full name:
Sarcoplasmic Reticulum Ca²⁺ ATPase
Function:
pumps Ca²⁺ from cytoplasm back into SR
This is:
the major mechanism causing relaxation
Important:
ATP-dependent
active transport
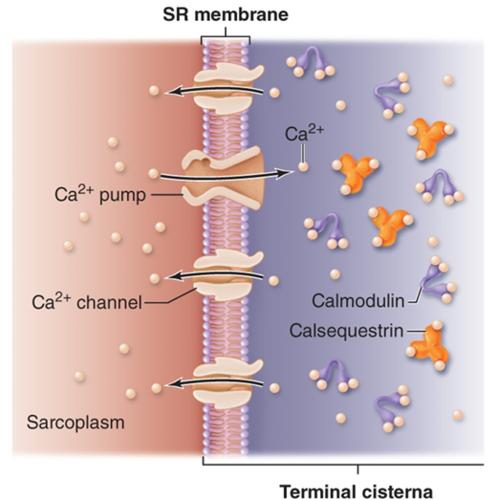
What Happens to Ca²⁺ Inside SR?
After Ca²⁺ enters SR:
it binds storage proteins:
calsequestrin
calreticulin
Purpose:
store large amounts of Ca²⁺ inside SR
Phospholamban & SERCA
Phospholamban
Regulates:
SERCA activity
Important point:
phosphorylation of phospholamban enhances SERCA function
Meaning:
more Ca²⁺ pumped into SR
relaxation becomes faster
Especially important in:
slow-twitch skeletal muscle fibers
Key Physiological Idea
Contraction
↑ cytoplasmic Ca²⁺
Relaxation
↓ cytoplasmic Ca²⁺
So:
relaxation depends on Ca²⁺ removal
High-Yield Final Exam Notes
Must Memorize
SERCA is the most important Ca²⁺ removal mechanism.
SERCA pumps Ca²⁺ back into SR.
SERCA requires ATP.
NCX and PMCA remove Ca²⁺ across sarcolemma.
Ca²⁺ stored in SR by calsequestrin and calreticulin.
Phosphorylation of phospholamban enhances SERCA activity.
Specialized Energy Stores in Muscle Cell
1. Phosphocreatine System
Immediate ATP reserve in muscle.
Creatine kinase transfers phosphate from phosphocreatine to ADP → ATP.
Very rapid ATP regeneration.
Effective for <10 seconds of intense activity.
2. Glycogen / Anaerobic Glycolysis
Glycogen = major stored energy source in skeletal muscle.
Glycogen → pyruvate → ATP.
Occurs by anaerobic metabolism (without O₂).
Can sustain muscle activity for ~1 minute without oxygen.
3. Oxidative Metabolism
Pyruvate further metabolized by oxidative metabolism.
Main long-term ATP source.
Requires oxygen.
ATP production depends on oxygen delivery to muscle.
High-Yield Points
Creatine kinase regenerates ATP rapidly.
Phosphocreatine supports short bursts of activity.
Glycogen supports anaerobic ATP production.
Oxidative metabolism is the major long-term ATP source.
Oxygen availability limits oxidative ATP generation.
Muscle Fatigue
Definition
Muscle fatigue =
inability to maintain desired power output during contraction.
Results in:
↓ force production
↓ velocity of shortening
Why Fatigue Occurs
Main causes:
↓ number of active cross-bridges
↓ force produced per cross-bridge
Important Features of Fatigued Muscle
Force declines earlier and more than shortening velocity.
Slower contraction and relaxation.
Fast movements become difficult.
Reason:
impaired Ca²⁺ release and reuptake from SR
Protective Role
Fatigue may protect muscle by:
reducing contraction rate and force
preventing cellular damage
Important Point
Muscle fatigue:
reversible with rest
Unlike muscle damage/weakness:
force generation remains impaired even after rest.
Types of Fatigue
1. Central Fatigue
Occurs in:
CNS
Causes:
altered sensory input
reduced excitatory input from brain/spinal cord
altered motor neuron excitability
2. Peripheral Fatigue
Occurs in:
muscle itself
Causes:
impaired muscle excitability
impaired Ca²⁺ release
Metabolic Causes of Fatigue
ATP depletion
lactic acid accumulation
glycogen depletion.
Muscle Fiber Types
Slow Fibers (Type I, Red Muscle)
Characteristics
Small muscle fibers
Innervated by small nerve fibers
Rich blood supply and many capillaries
Many mitochondria
High myoglobin content
Metabolism
Mainly:
oxidative metabolism
Slow but efficient ATP production
Functional Features
Fatigue resistant
Good for prolonged activity/endurance
Why Called “Red Muscle”?
High myoglobin content gives red color.
Myoglobin stores and transports oxygen to mitochondria.
Fast Fibers (Type II, White Muscle)
Characteristics
Large muscle fibers
Strong contractions
Extensive SR for rapid Ca²⁺ release
High glycolytic enzyme content
Fewer mitochondria
Less blood supply
Low myoglobin content
Metabolism
Mainly:
glycolytic / anaerobic metabolism
Rapid ATP production
Functional Features
Powerful rapid contractions
Fatigue easily
Why Called “White Muscle”?
Low myoglobin content gives pale/white appearance.
Cardiac Myocyte Structure
General Features
Cardiac myocytes are:
shorter than skeletal muscle fibers
branched
interconnected end-to-end
Intercalated Disks
Cardiac cells connected by:
intercalated disks
Contain:
desmosomes
gap junctions
Functions of Intercalated Disks
Desmosomes
mechanical attachment between cells
prevent separation during contraction
Gap Junctions
allow electrical current to pass between cells
enable rapid spread of action potentials
Functional Syncytium
Cardiac muscle acts as:
mechanical and electrical syncytium
Meaning:
cells contract together as one unit
electrical activity spreads rapidly through heart
Important Timing
AP from SA node spreads through heart in:
~0.22 sec
Cardiac muscle contraction lasts:
~0.3 sec
Sarcolemma Structures
Includes:
T tubules
terminal cisternae
sarcoplasmic reticulum (SR)
These structures participate in:
excitation–contraction coupling
High-Yield Points
Cardiac myocytes are branched and interconnected.
Intercalated disks contain desmosomes and gap junctions.
Desmosomes = mechanical coupling.
Gap junctions = electrical coupling.
Cardiac muscle functions as a syncytium.
AP spreads through heart rapidly from SA node.
Phospholamban Effect on Heart Activity
Phospholamban
Regulatory protein in SR membrane.
Regulates:
SERCA pump
Normal Function
Phospholamban normally:
inhibits SERCA
So:
less Ca²⁺ pumped back into SR
slower relaxation
Phosphorylation of Phospholamban
Stimulated by:
β₁-adrenergic stimulation
Mechanism:
β₁ stimulation
→ PKA activation
→ phospholamban phosphorylation
Effect of Phosphorylation
Phosphorylation removes phospholamban inhibition on SERCA.
Result:
SERCA activity increases
So:
Ca²⁺ reuptake into SR becomes faster
more Ca²⁺ stored in SR
Physiological Effects
1. Faster Relaxation
Because cytoplasmic Ca²⁺ decreases faster.
This is:
positive lusitropic effect
2. Stronger Next Contraction
Because more Ca²⁺ stored in SR for next beat.
This is:
positive inotropic effect
High-Yield Points
Phospholamban inhibits SERCA.
Phosphorylation disinhibits SERCA.
β₁ stimulation → PKA → phospholamban phosphorylation.
Increased SERCA activity → faster relaxation.
More SR Ca²⁺ storage → stronger contraction.
Action Potential Generation in Smooth Muscle
General Principle
Stimuli produce:
graded changes in membrane potential (Vm)
These changes may be:
depolarization
hyperpolarization
Summation
Graded potentials can summate:
temporally
spatially
If depolarization reaches threshold:
action potential occurs
Types of Stimuli
1. Chemical Stimuli
circulating hormones
local humoral factors
2. Mechanical Stimuli
stretching of smooth muscle cell
Spontaneous Electrical Activity
Some smooth muscle cells generate spontaneous activity.
Two important mechanisms:
1. Pacemaker Cells
Interstitial cells of Cajal
Function:
initiate rhythmic contractions
especially in GI tract
2. Slow Waves
Slow waves =
spontaneous oscillations in membrane potential
Important:
do not always produce APs
if threshold reached → spike potentials/APs occur
Mechanism of Slow Waves
Voltage-gated Ca²⁺ channels active at resting Vm
→ gradual depolarization and Ca²⁺ influx
Then:
Ca²⁺-dependent K⁺ channels open
→ hyperpolarization
This cycle repeats rhythmically.
Effects of Autonomic Input
Depolarization / Excitation
Caused by:
stretch
acetylcholine
parasympathetic stimulation
→ increases AP firing and contraction
Hyperpolarization / Inhibition
Caused by:
norepinephrine
sympathetic stimulation
→ decreases excitability and contraction.