Lecture 7: Regulation, Enzymes and Rate Limiting Steps
Learning Outcomes:
Describe the chemical features of ATP which make it ideal for use as an energy currency
Explain the concept of energy charge with reference to the concentration of adenine nucleotides
Review how a small change in ATP concentration is translated into a large relative change in AMP concentration
Identify the most likely control points in metabolic pathways
Interpret enzyme kinetic parameters to identify potential rate limiting steps
Describe the properties of rate limiting steps
Review the major ways in which enzyme activity can be changed
List the key rate limiting steps in the major pathways of catabolism
Provide an overview of the regulation of phosphofructokinase
Provide an overview of the regulation of hexokinase
Using an example, illustrate how control motifs act synergistically to regulate pathways
Using an example, show how enzymes are controlled by reversible phosphorylation
Recognise that rate limiting steps can change with circumstances
Explain the principles of reciprocal regulation of pathways
Cells are always checking how much energy they have – need to keep ATP at 5mM
Large changes in ATP are not desirable
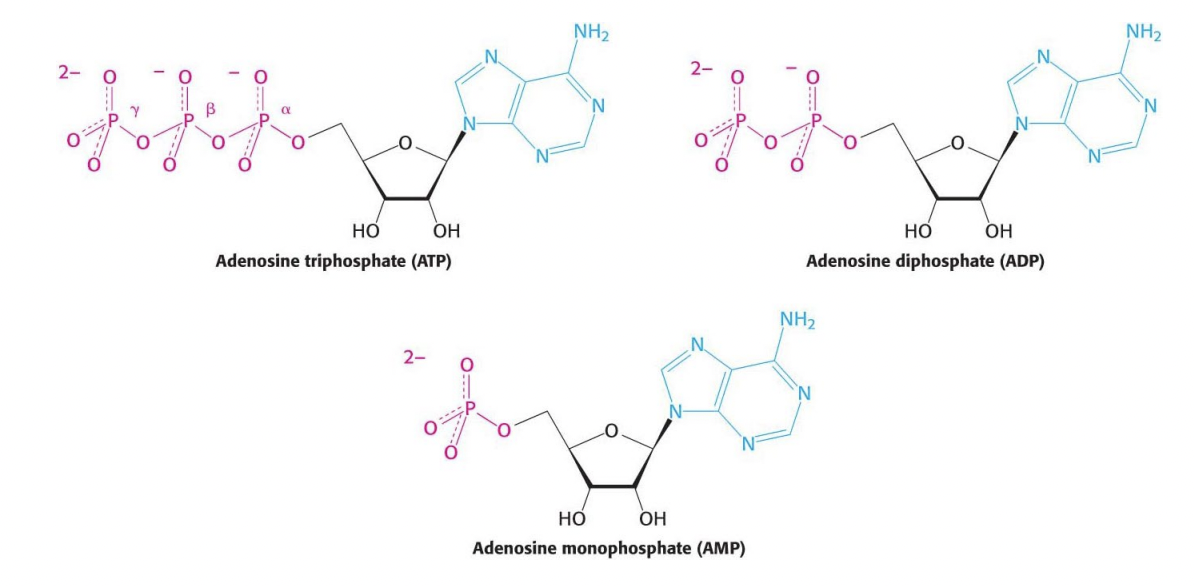
ATP is not the most high energy molecule in our cells:
Higher energy molecules can transfer their phosphate group onto ADP – substrate level phosphorylation
Creatine phosphate – when you want to start sprinting quickly – burst of ATP that wasn’t stored in cells
Passes phosphate onto ADP as it is in a higher energy state
1,3-Bisphosphoglycerate (second half of glycolysis – energy return phase)
3 carbon sugar phosphate and adding another phosphate to it
Phosphenolpyruvate (PEP)
Also at the end of glycolysis
Very high energy phosphate bond
Above 3 molecules are instant reserves of high energy phosphate – can only supply for a few seconds
ATP:
energy released when any terminal phosphates are hydrolysed
ATP to ADP releases energy
ATP to AMP releases energy
Instant ATP:
Adenylate kinase (enzyme)
Take 2ADP and create an ATP and AMP
Energy Charge = measure of how much energy is in a cell – ratio of adenine nucleotide concentrations
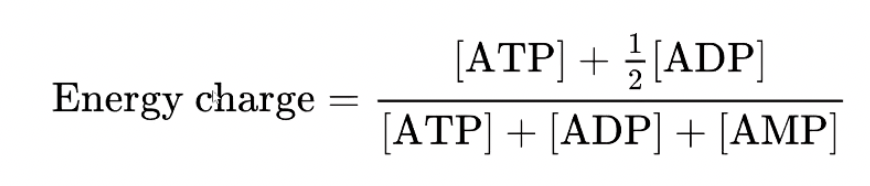
AMP only appears on the bottom so large changes to AMP will reduce the energy charge of a cell significantly
Concentration of ATP in cells depends on type of cell and the organism – not always 5mM
BUT [ATP] > [ADP] > > [AMP]
AMP concentration is always very low
Energy Charge and AMP:
AMP is the most sensitive watch on energy charge
Has the largest relative change when all energy is depleted in cells
Easy for cells to watch AMP to signal for energy charge getting low

Which enzymes are controlled:
Slowest enzyme in the pathway determines the overall speed
Rate-limiting steps
Flux generating steps
Flux = flow of molecules on a pathway
Don’t need to control every enzyme involved
These enzymes being controlled are usually low Km
Don’t have a high reaction velocity
Usually working at substrate concentrations that are much higher than their Km value
At high substrate concentrations, minor changes in substrate concentrations will not affect the rate of the reaction
Doubling or halving the substrate concentration isn’t even going to effect the rate
The only time substrate concentration concentration impacts reaction rate is when it reaches Km value
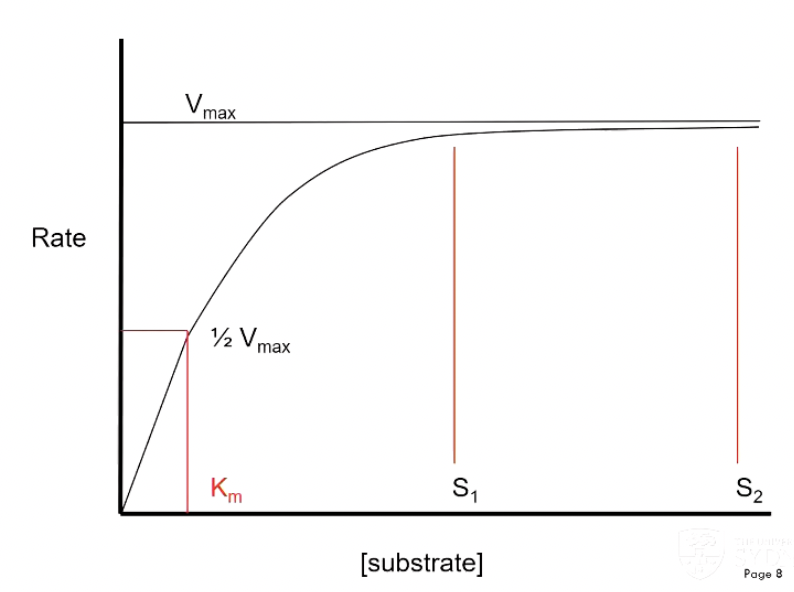
Km = substrate concentration at which the enzyme is catalysing the reaction at half the maximum reaction velocity
When you are near Km, small changes in substrate concentration is going to effect the rate of reaction
x10 or x50 Km – changing substrate concentration will not effect the reaction velocity
Properties of RLS (rate limiting steps):
Irreversible
Need alternative enzymes to go back
Not equilibrium – not effected by substrate or product concentrations
Committed steps – once you’ve gone past a point, theres no way to get back without another enzyme
Saturated with substrate
Low kM or [substrate] > Km
Working at Vmax
e.g. Peak hour at train station barriers – barrier = enzyme working at maximum velocity – lots of people (substrate)
Increasing or decreasing passengers isn’t going to change the rate at which people go through the gate
Controlling one gate (enzyme) impacts how long all the other people take to get home or to their work (substrate)
3 major ways to regulate RLS:
Change the intrinsic activity of the step
Make the rate limiting enzyme go faster or slower
Make ticket reading/gate opening happen faster
Make more gates open
Turn the rate limiting enzyme on/off or make it work the other way
Switch gates from off to on
Or change the direction from in to out
Make and destroy gates according to need
Increase the rate of transcription/translation of the rate limiting step or change its rate of degradation
Bring in a new set of gates when you need them
Rate Limiting Factors in Catabolism:
Changes under different conditions
Fed, starved, resting, exercising, nutrients consumed etc.
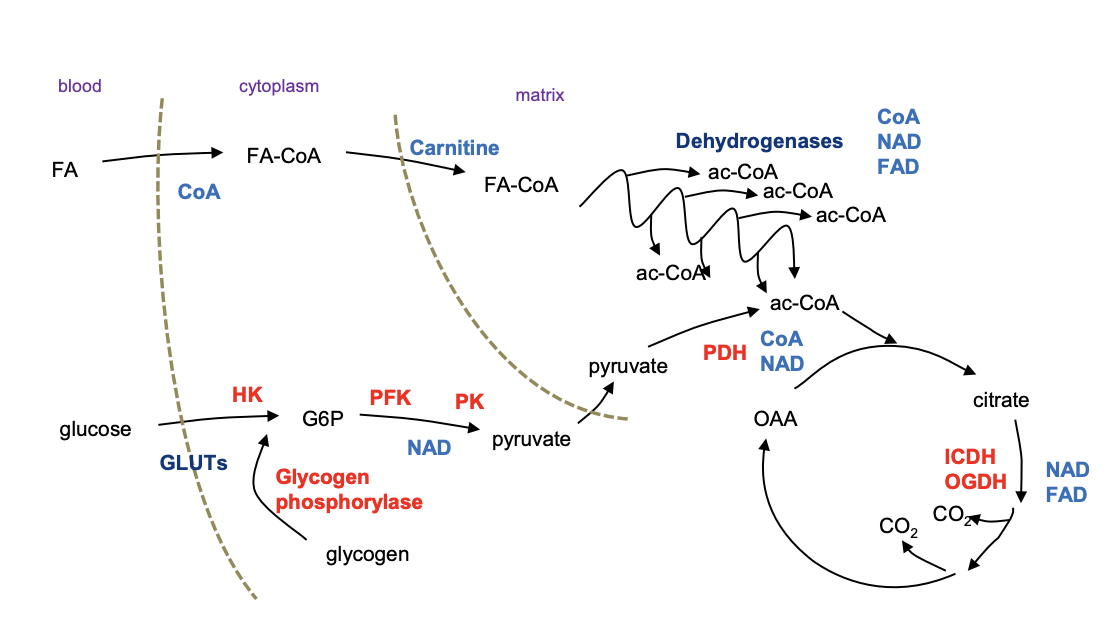
Don’t regulate every step:
Can regulate enzyme activity or the availability of cofactors
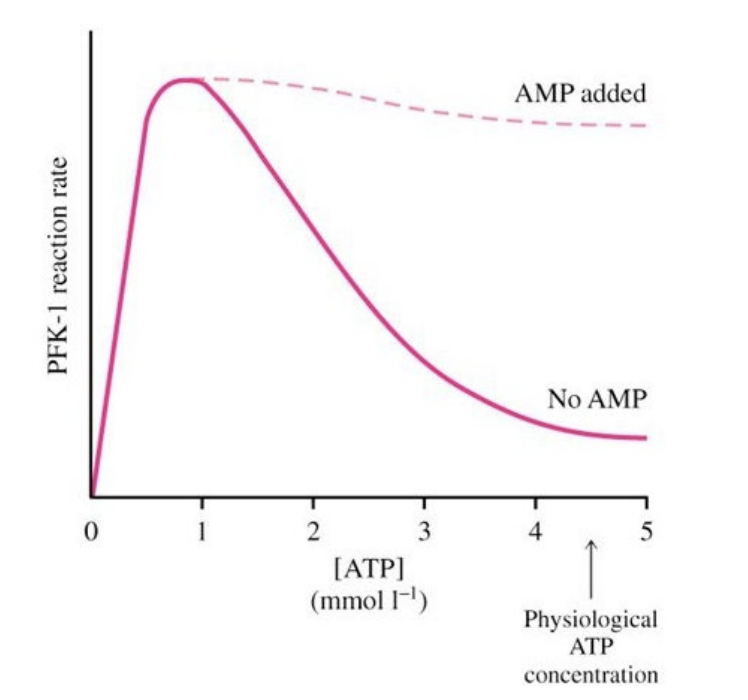
PFK Enzyme – phosphofructokinase
Does not like a high concentration of its’ substrate, ATP
Does not have a high affinity for ATP
Activity of PFK is low at 5 mmol (the baseline concentration of ATP)
Once AMP is added, activity is increased dramatically
Through allosteric activation:
Activator binds allosterically (at a site away from the active site) on the enzyme, activates it, changes the shape of it to give it a higher affinity for its’ substrate
This is how AMP impacts phosphofructokinase
PFK also binds citrate allosterically, changes the shape of the active site to switch it off
Citrate inhibits PFK
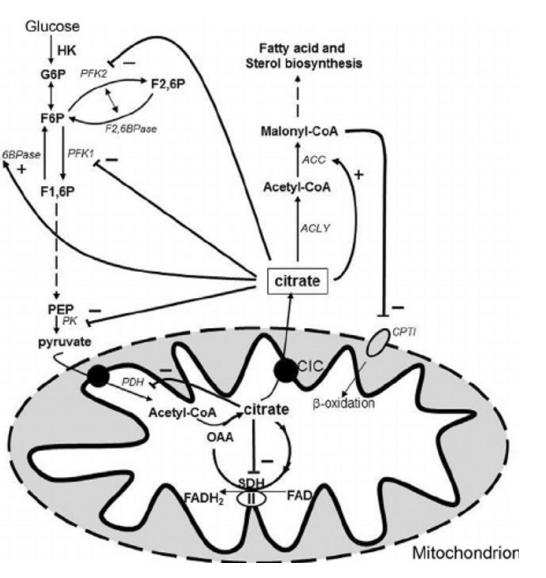
Lots of citrate = lots of acetyl-CoA
Citrate can go back out into the cytoplasm and tell PFK to switch glycolysis off – allosteric inhibition
Hexokinase Enzyme – Feedback Inhibition
First step in glycolysis, traps glucose in the cytoplasm by phosphorylating it to creat GP6 (adds a phosphate group) using ATP
Inhibition by not using the product (GP6) prevents excessive trapping
Prevents ATP waste
Allows glucose to not go back out the cell
As GP6 builds up in the cell, negative feedback inhibits hexokinase
If GP6 is not being used, more glucose is not being trapped
If GP6 is used in the next step o glycolysis, inhibition is relieved and more glucose is trapped in the cytoplasm.
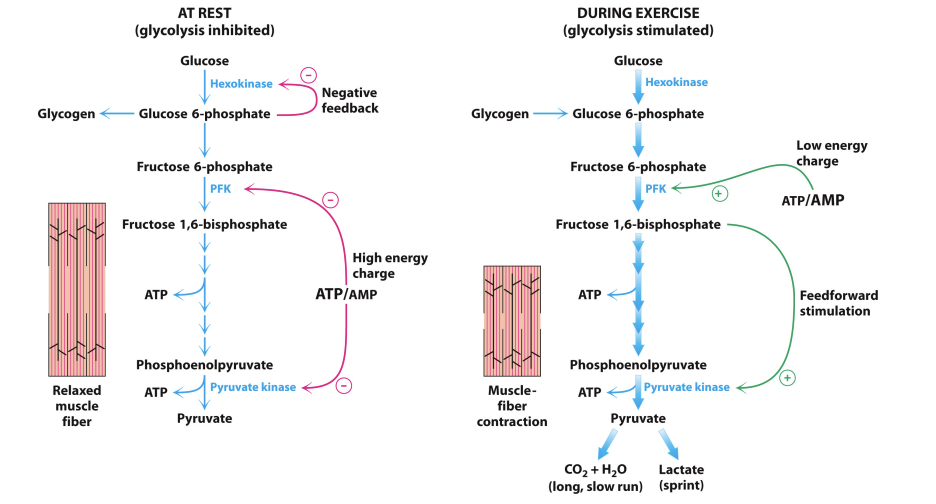
Example – Glycolysis in Exercise:
At rest (Glycolysis inhibited):
Glucose → GP6 but since there is no exercise, GP6 build up occurs
Negative feedback inhibits hexokinase from trapping more glucose in the cytoplasm
Energy charge is quite high while resting – lots of ATP, not much AMP
High energy charge acts of PFK (phosphofructokinase) and is getting switched off due to minimal AMP – it is not very attracted to ATP
Pyruvate kinase is also being switched off (at the end of glycolysis) by the high energy charge – not converting much ATP
During Exercise (glycolysis stimulates):
Glucose → GP6 but GP6 is being used so inhibition of hexokinase is relieved
Hexokinase can start trapping more glucose as it comes into the cell
Energy charge is now higher in AMP than ATP – overall low energy charge
PFK is becoming switched on and has a higher affinity binding to ATP due to increased levels of AMP – AMP allosterically activates PFK to switch on glycolysis
Feedforward stimulation from fructose 1,6-bisphosphate to pyruvate kinase to increase activity
feedforward stimulation would be like people at back of queue shouting for person at the front to hurry up
In high FAO (fatty acid oxidation),
High [citrate] = lots of acetyl-coa entering the krebs cycle
If citrate goes out into cytoplasm, it will switch of glycolysis by signalling PFK and pyruvate kinase that there are enough Kreb cycle intermediates
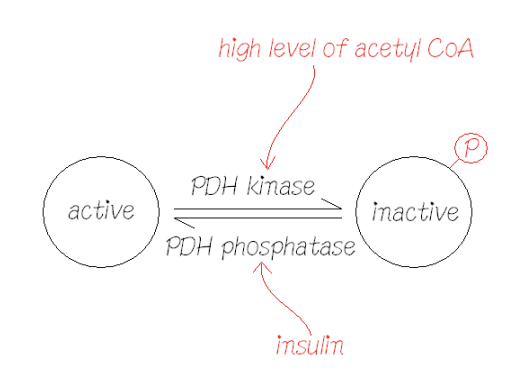
PDH (Pyruvate dehydrogenase) – covalent modification NOT allosteric inhibition/activation:
Inactivated entirely by phosphorylation
Covalent attachment of phosphate, catalysed by PDH kinase
Total amount of enzyme doesn’t change – just the ratio of phosphorylated to dephosphorylated
Reactivation by phosphate PDH phosphotase – release of phosphate = totally on
PDH activity a balance between kinase and phosphate
Kinases use ATP to add a phosphate to their substrate
High level of acetyl coa activates PDH kinase which adds a phosphate group to inactivate PDH
Insulin acts on PDH phosphatase to remove the phosphate from PDH to activate it again
The RLS in Glycolysis, β-oxidation, Krebs?
Could be availability of substrate (generally not)
Cell membrane transport and trapping
Mitochondria transport
Carintine availability
Oxidative capacity
Total activity of enzymes
Supply of oxygen (Electron transport chain – oxygen as the terminal electron acceptor)
It will depend on the circumstances
Overarching is cofactor availability (NAD+, FAD, CoA) an AMP level
Which itself is dictated by demand for ATP
Catabolism vs Anabolism:
Glycolysis vs gluconeogenisis
β-oxidation vs FA Synthesis
When one pathway is stimulated, the opposing pathway is inhibited
When they both occur at the same time it is a futile cycle
Not going to be breaking something while you’re also making it
Not always true – e.g. specialised tissues
Citrate as an Example:
Citrate is the first product in the krebs cycle
When citrate makes its way to the cytoplasm because there are enough Krebs cycle intermediates, citrate switches off glycolysis and on gluconeogenisis
Citrate switches of β-oxidation and on FA Synthesis
Balance of catabolic and anabolic pathways