lecture 12: The ER as a portal and stress sensor
To the golgi
Summary so far: 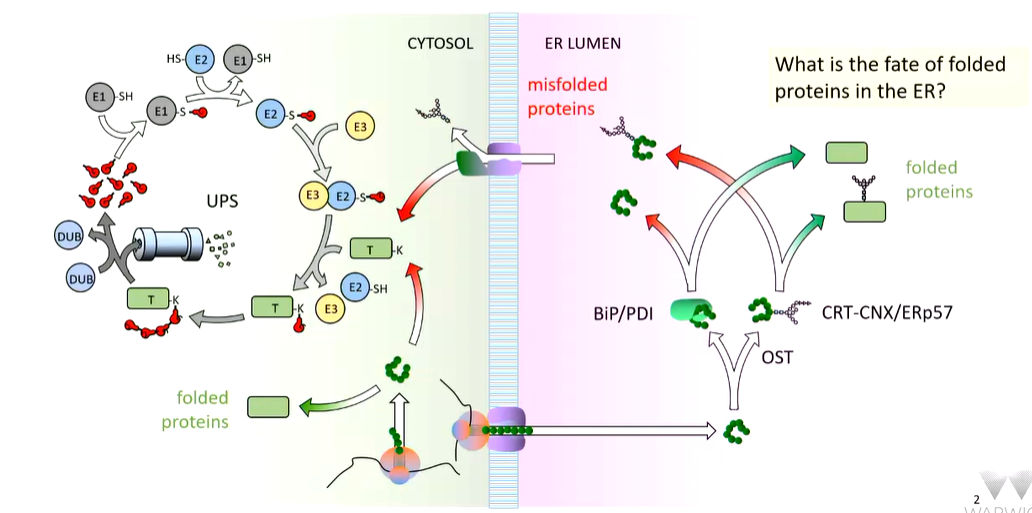
roadmap of intracellular trafficking:
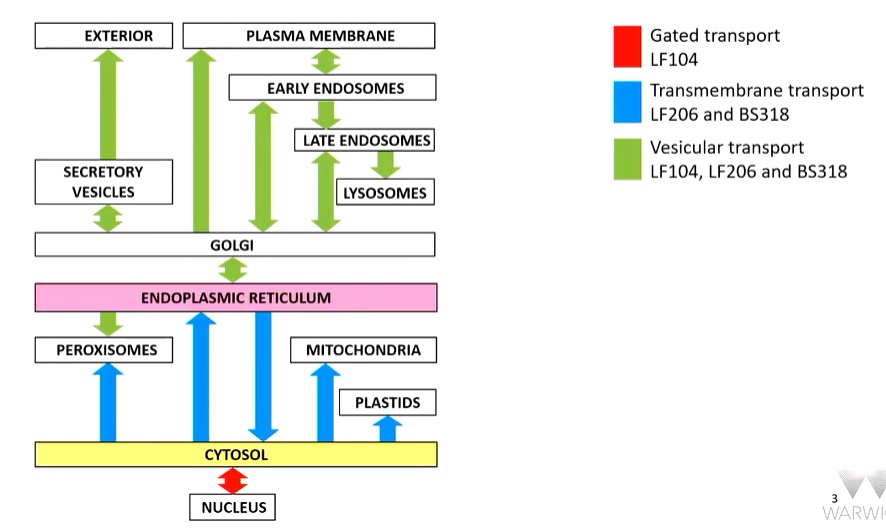
There are multiple destinations that can be accessed from the ER.
There are 5 major interdependent strategies (selective transport) for how some proteins are retained in the ER and how some pass onto the secretory pathway:

bulk flow - just the flow of proteins through the system with no cargo capture
These 5 strategies maintain ER homeostasis and reduce cellular stress
Cargo capture and anterograde transport
This is transport from the ER to the golgi. The cis golgi is the side facing the ER
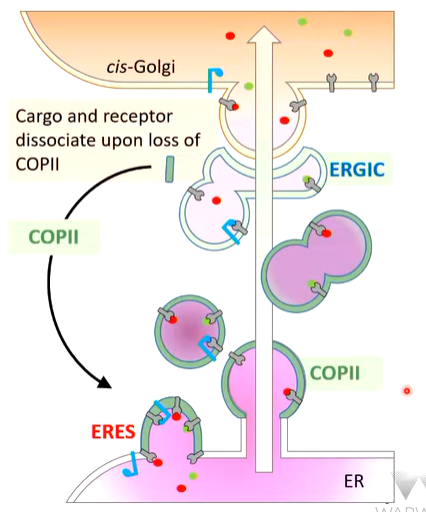
There is a directionality from the ER to the cis golgi and starts at orgellar buds called ERES (ER exit sites). The bud (ring) is coated with a set of COP II proteins and this organises anterograde transport. The ERES starts to swell and is nipped off. COP II coats capture cytosolic sites of transmembrane receptors - receptors for soluble materials that piece the membranes and are captures as well as transmembrane receptors. The ERES buds off and moves towards the golgi
On the way, the buds start to fuse and cause the formation of ERGIC (ER golgi intermediate compartment). As they fuse, the COP II coats are ejected and recycled for the next round. The ERGIC vesicles fuse with the cis golgi and a change in pH allows the vesicles to let go of their cargo. The cargo and receptor dissociate upon the loss of COP II
Bulk flow and retention
Bulk flow is the transport of proteins through a system in an anterograde manner with no receptors. They enter the ERES sites in a receptor independent manner that bud off ad are transported in the forward flow of membranes and vesicles
This is true for some membrane proteins and soluble proteins
An advantage for this is for biotech - if a plant protein is taken and expressed in yeast there is a lack in species specific transport recognition signals. In biotech, if a transformed yeast is directed into the ER, it will get to the outside of the cell in a receptorless manner. If it is a soluble proteins, it is easy to purify
retention
This is another mechanism where some proteins are selectively excluded from COP II vesicles - no access route for them to move forward. 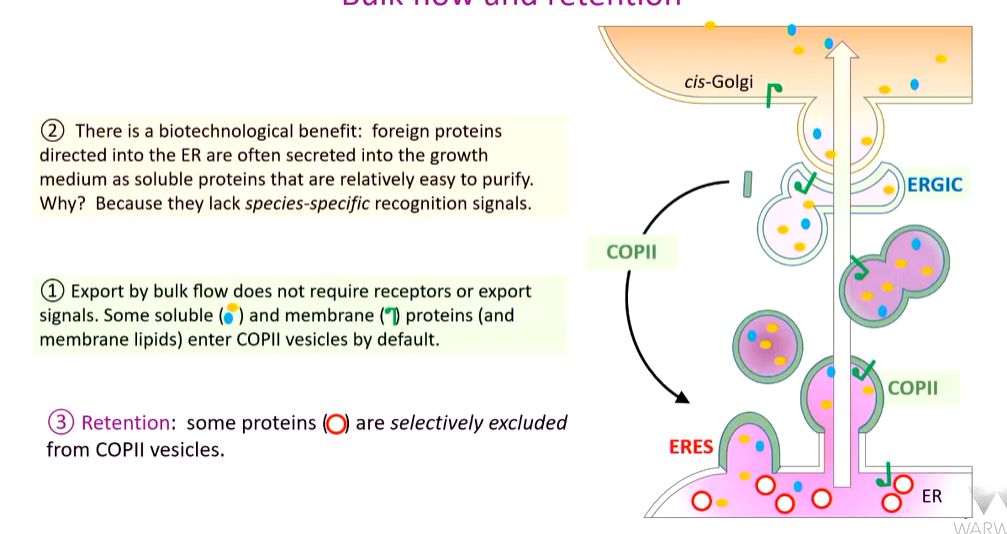
Retrieval and retrograde transport (COP I)
Any cargo receptors that have trafficked the vesicles to the golgi must be retrieved as well as proteins and receptors from ERGIC compartments
COPI coated vesicles and captures receptors that are retrieved and is also itself recycled. The ERGI compartment is a combination of both COPII and COPI proteins. This allows a nuanced distinction of destinations
Retrieved membrane proteins Possess a C terminal dilysine motif (KKXXX) or a close variant (EKEKSD). This is because some other proteins will end up in the wrong place such as membrane proteins such as OST that will only work in the ER. This has a modified dilysine motif - 2 positive charges make a good candidate for retrieval
Retrieved soluble proteins have a c terminal KDEL motif that is HDEL in plants 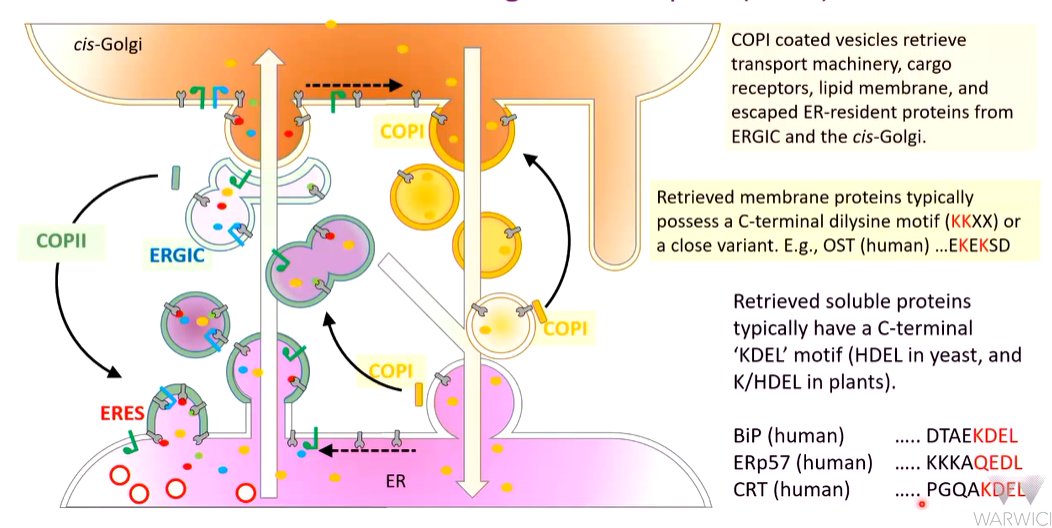
If vesicles in an antegrade fashion are sent off that means that membranes are depleted of their lipids. There is an extra transport tep that is governed by small g proteins such as Rabt. This organises the return via ;og tubule elements of membrane proteins that have to be returned to their place of origin. This is independent of COPI.
Shiga toxin binds a lipid receptor on the cell surface utilizes the Rab6 pathway. Lipids from the surface of the cell can be retrieved

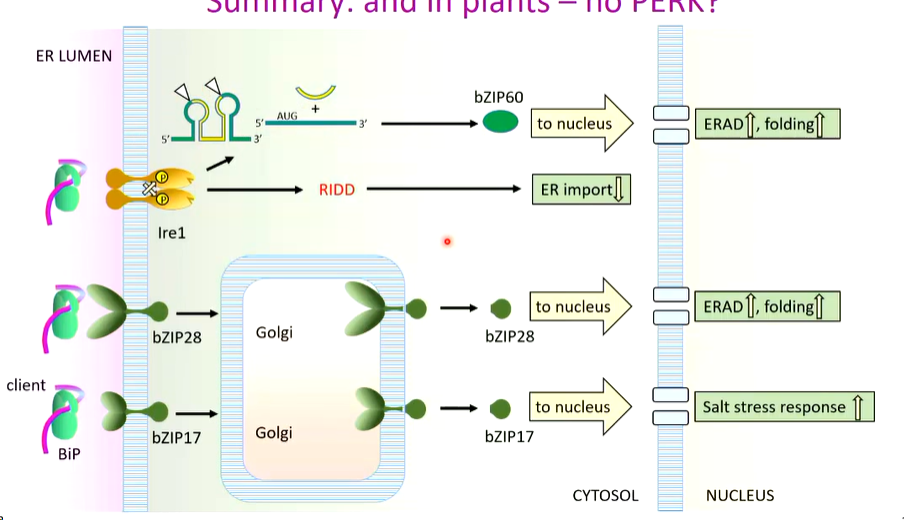
IRE senses stress
There are multiple stresses that cause damage to proteins:
abiotic stresses
heat stress, osmotic stress, high light intensity
biotic stresses
infection
stress related hormones

The ER can act as a stress sensor and if the folding protein capacity is breached, it can signal and sets into motion pathways to reduce the stress
This is call the unfolded protein response (UPR) and once this is triggered, physiological responses take place
there are three stress sensors:
IRE1
universal in eukaryotes
PERK
in animals and fungi
ATF6
animals and fungi
bZIP28 and bZIP17 in plants
Between them, these systems monitor ER stress by counting how much proteins are unfolded and adjust the folding protein capacity of the ER to cope
IRE1
This is universal
When the ER is unstressed, BiP is bound to IRE1 in the cytosol transmembranal to Bip in the ER lumen. Bip Maintains IRE1 as soluble and keeps it inactive.
In Low ER stress, Bip detects the misfolded proteins and leaves. IRE1 dimerises and binds unfolded proteins. In increasing ER stress, there is oligomerization if IRE1 which are activated.
The bringing together of these molecules allows auto transphosphorylation. The cytosolic domains activate each other and depending on the level of stress, the more IRE1 is phosphorylated.
The activation of IRE1 is a kinase as well as an RNAse that recognises the unusual highly conserved cytosolic RNA that has two stem and loop structures. The IRE1 cleaves at the triangles 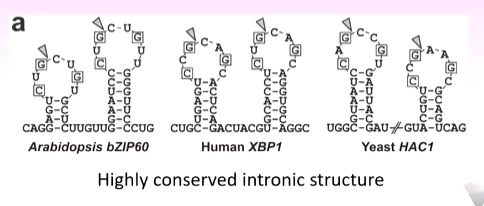
There are two exons that are spliced together leaving a cytosolic intron. The exons creates an open reading frame that is translated to make a transcription factor that turns on response pathways that upregulate ERAD, folding rate by increasing chaperones and trafficking rate
splicing mechanism
In IRE1, it is more cut and paste that is specific to the stress response. 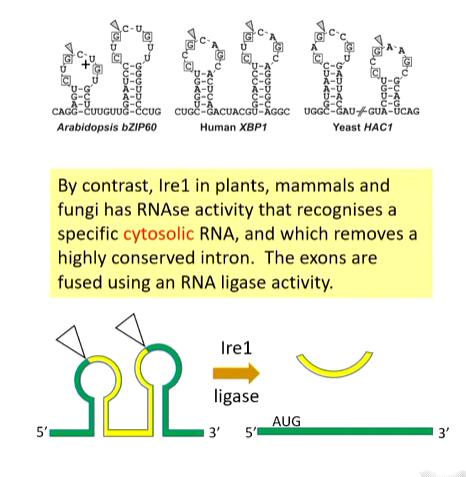
How do we measure stress in ER experimentally
DTT is a reducing reagent that breaks disulfide bonds. If cells are treated with DTT, their ER proteins are destabilized so DTT stimulates stress.
TG (thapsigargin) blocks sarco/endoplasmic reticulum Ca ATPase (Serca) pumps. This means that Ca ions are not pumped to the ER so it cannot store it. This leads to Er stress as this is also important for folding
Heat shock destabilizes and unfolds some proteins
Monitoring IRE1 mediated splicing as a measure of ER stress
Adding ATT stimulates ER stress, Hac1p is expressed from spliced Hac1 mRNA (yeast version of IRE1). In mammals, activated IRE1 spliced XBP1 mRNA. TG leads to ER stress and XBP1s protein is expressed from spliced XBP1 mRNA 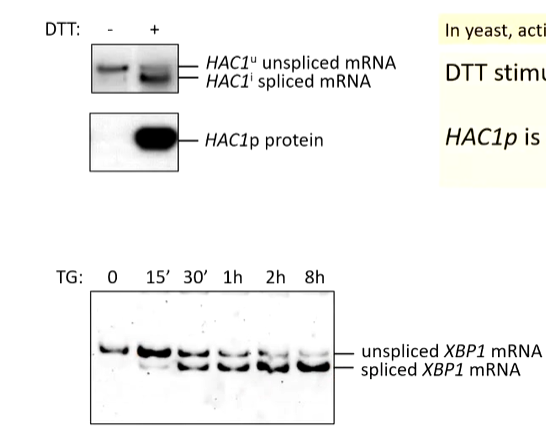

IRE1 RNase activity
IRE1 also has general RNAse activity (RIDD). RIDD is regulated IRE1 dependant decay of mRNA. Any mRNA close to the ER membrane are degraded non specifically. Cytosolic protein production continues but there is less transport into the ER which reduced stress
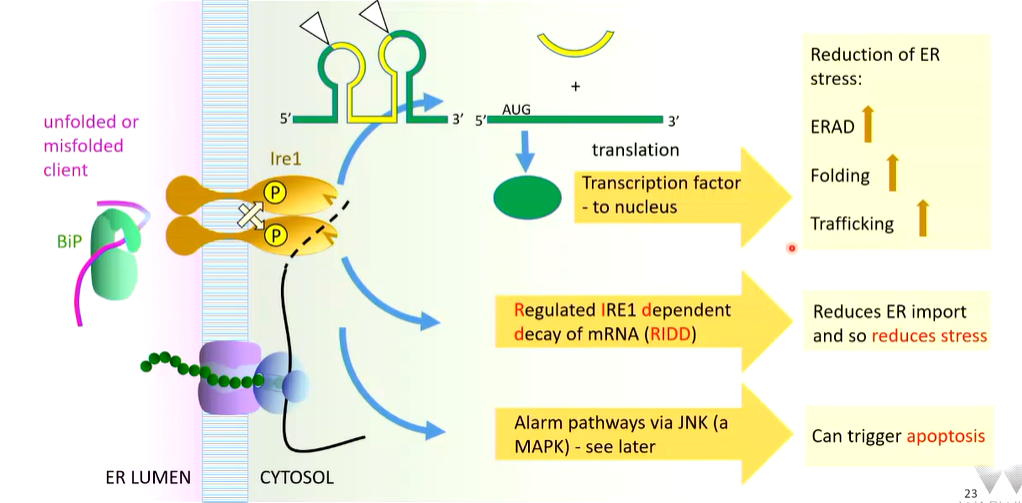
Other stress sensors
PERK and IRE1

Mechanism
PERK interacts with BiP in the same way as IRE1 and is also inactivated by it. In ER stress, it acts a similar way to IRE1 where unfolded proteins allows for the multimerization of PERK that leads to auto transphosphorylation thata ctivates PERK.
There is phosphorylation of eLF2a which is required for translation of all proteins (almost). When it is phosphorylated it is inactivated so the elongation step does not occur protein production does not occur. This reduces stress a
Some RNAs are preferentially translated. There is stimulation of specific translation of some gene such as ATF3 that makes a transcription factor transported to the nucleus. This results in the expression of CHOP that leads to apoptosis. 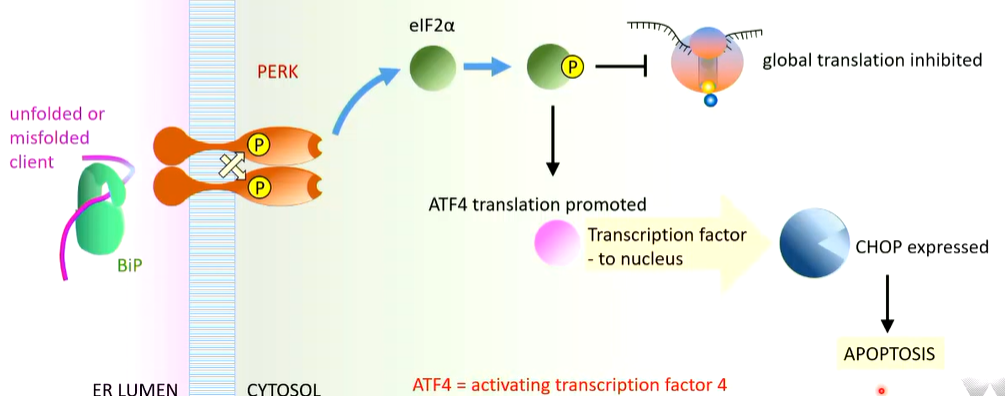
ATP6 mechanism
In no ER stress it is bound to BiP like IRE1 and in the same way, it is inactivated. ER stress means bip is competed away.
BiP has a KDEL sequence meaning it is reteried easily so ATP6 is transported into the golgi as it has lost the KDEL sequence. There are specialised proteases such as single peptidases in the ER but as you go through the secretory pathway, you start seeing more. IN the golgi, there is one that recognises ATF6 that cleaves it and leaves a fragment called ATF6F that acts as a transcription factor that upregulates Bip and other ER chaperones and ERAD,
summary
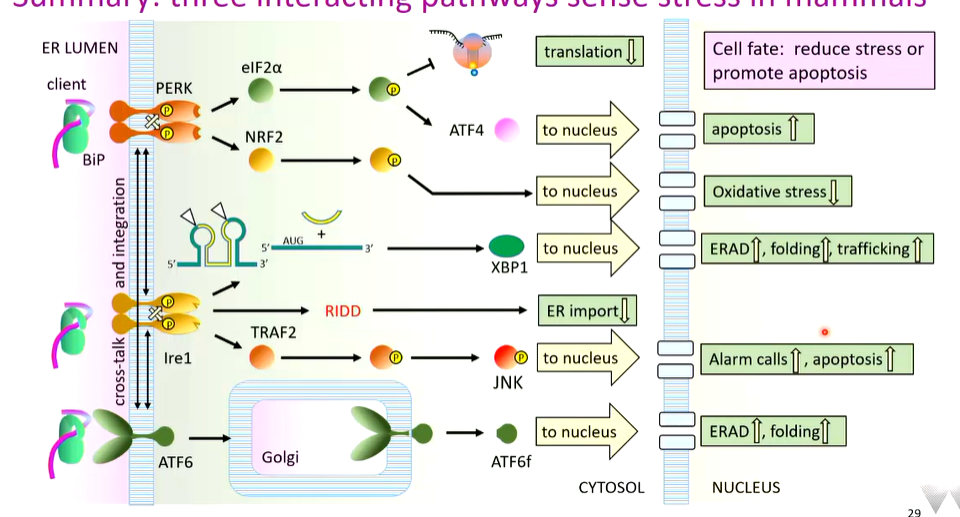
take home - stress responses are matched to type and quantity of stress so there is appropriate reducing or death
if youre a plant there is no PERK