Lecture 12
Conjugation process
A pilus is a long hollow protein tube made of repeating protein subunits called pilin which are encoded by plasmid genes
The donor bacterium extends a pilus to contact the recipient
The pilus pulls the cells together and a stable mating bridge forms
A copy of the plasmid DNA moves through the hollow channel into the recipient
The plasmid is usually copied during transfer so that the donor keeps its plasmid as well
Transfer begins at the origin of transfer (oriT)
Relaxase is a conjugation enzyme that works with other transfer proteins (tra genes) to form the relaxosome
Relaxase recognises oriT and nicks phosphodiester bonds at one DNA strand which creates a free single-stranded DNA end
This generates a free 5’ end and a transferable single strand that is transferred into the recipient
DNA movement through the membrane pore and conjugation pilus requires energy so conjugation is ATP-dependent
As the DNA moves, both donor and recipient simultaneously synthesise complementary strands
The recipient cell receives single-stranded plasmid DNA and generates the complementary strand (lagging strand synthesis)
The donor cell replaces the strand being exported so synthesises the leading strand
Formation of HFR strains
Occurs when the F plasmid integrates into the bacterial chromosome (High Frequency of Recombination)
The F plasmid and chromosome both contain insertion sequences and these shared DNA sequences provide sequence homology
Because the IS elements are similar, recombination can occur and the plasmid inserts itself into the chromosome
The plasmid can no longer exists independently but the tra genes still function, so the bacterium can still conjugate
It is known as HFR as during conjugation, transfer starts at oriT inside the integrated F factor but chromosomal DNA adjacent to the F plasmid also gets transferred
This greatly increases chromosomal gene transfer
So the HFR cell transfers part of the integrated F plasmid and nearby chromosomal genes into a F⁻ recipient
It is important to note that the entire chromosome is usually too large to fully transfer before the mating bridge breaks so recipient often receives chromosomal genes but not the complete F factor, therefore recipient usually remains F⁻
F⁺ cell → has free F plasmid and can conjugate
F⁻ cell → lacks F plasmid, recipient only
F′ (F-prime) cell → occurs when the integrated F plasmid excises incorrectly from the chromosome, plasmid contains F plasmid DNA and chromosomal bacterial genes
Hfr cell → F plasmid integrated into chromosome so transfers chromosomal genes efficiently
Gene mapping
To map genes, you can use specific genotypes as an appropriate complementary/comparator genotype
In an Hfr strain, F plasmid is integrated into the chromosome, transfer starts at oriT so chromosomal genes are in order
Therefore, genes closer to oriT enter the recipient earlier whilst genes further away enter later
This allows for determination of gene order and approximate distances between genes
Interrupted mating experiments:
A Hfr donor mates with an F⁻ recipient and DNA transfer starts from oriT and proceeds linearly through the chromosome
Researchers stop conjugation at different times, 5, 10, 20 minutes to physically stop DNA transfer
Transferred DNA can recombine into the recipient chromosome and the recipient acquires new traits
Count the amount of transconjugants that gained a specific phenotype to reveal when a gene entered conjugation
Example using lacZ:
Recipient cell is lacZ⁻ and streptomycin resistant and the donor Hfr cell is lacZ⁺ and streptomycin sensitive
After mating, cells are plated on streptomycin and lactose as the sole carbon source
Streptomycin kills the donor cells and only the recipient-derived cells survive in a process called counterselection
Only transconjugants that have received the lacZ gene from donor will be able to grow on the lactose medium
Number of colonies obtained at different time intervals reflects distance of lacZ from the integrated F plasmid in donor
Cloning vectors
Plasmids or genetic constructs that accept inserted DNA fragments and are capable of replicating inside a host cell
Cloning vectors were originally derived from natural bacterial plasmids, bacteriophages or transposons and then genetically engineered for laboratory use
Majority are small, high-copy plasmids based on the ColE1 plasmid
Small plasmids are easier to manipulate, isolate and insert DNA into and high copy number plasmids have lots of plasmid DNA which leads to easier purification and stronger gene amplification
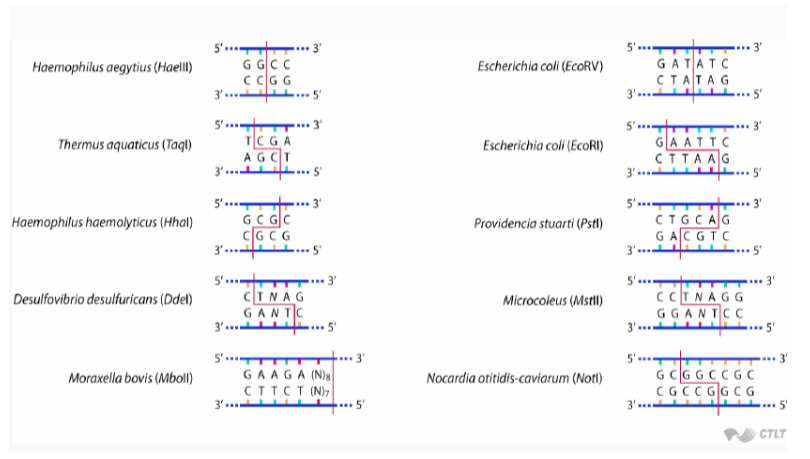
Most restriction enzymes recognise short sequences that are usually 4-6 bp long e.g. EcoRI recognises GAATTC
Palindromic sequences are sequences that read the same in the 5’ → 3’ direction on both strands
This is important as symmetry is common in restriction sites and restriction enzymes recognise these sequences as areas to cut
Restriction enzymes can either produce staggered, sticky ends that create short, single-stranded overhangs or straight, blunt cuts across both strands
Sticky ends are useful as they improve insertion efficiency and help orientate DNA fragments
Process:
A plasmid vector is cut using a restriction enzyme which opens the circular plasmid
The same restriction enzyme cuts the foreign DNA fragment to create sticky ends
The sticky ends naturally base pair which temporarily holds the DNA together
DNA ligase forms phosphodiester bonds to permanently join the plasmid DNA and inserted foreign DNA to form a recombinant plasmid
The original cloning vector: pBR322
Constructed by Rodriguez & Boyer (1977) and combined selectable markers, restriction sites and stable replication
Origin of replication was derived from the ColE1 plasmid to allow for plasmid replication in E.coli and a relatively high copy number
Contains two resistance genes: bla and tet to act as selectable markers
bla encodes beta-lactamase which gives ampicillin resistance so destroys beta-lactam antibiotics
tet provides tetracycline resistance via an efflux mechanism
Contains unique restriction enzyme sites such as EcoRI, HindIII, BamHI and PstI
Insertion at Pstl site:
Lies inside the bla (AmpR) gene
Foreign DNA insertion disrupts beta-lactamase production which results in the plasmid becoming ampicillin-sensitive but still tet-resistant
First transform bacteria with plasmids and then plate on tetracycline medium so only cells with the plasmid survive
Replica plate onto ampicillin medium and observe the colonies
Colonies that grow on tetracycline but not ampicillin likely contain recombinant plasmid because the tet gene still works but the bla gene is disrupted by DNA insertion
Insertion at BamHI site:
Lies inside the tet gene
Foreign DNA disrupts tetracycline resistance which results in tet-sensitive and ampicillin resistant plasmid
Plate on ampicillin and only plasmid containing bacteria survive, replica plate on tetracycline
Colonies that grow on ampicillin but not tetracycline contain recombinant plasmids because the tet gene was disrupted
Screening methods
Colony PCR allows scientists to pick colonies, amplify inserted DNA and confirm insert presence/size
PCR or sequencing verifying determines correct orientation as the insert can go in forward or reverse
Northern and Western blot can be used to test whether inserted gene is transcribed (RNA) or translated (protein) respectively
A modern cloning vector: pUC19
Small, high-copy plasmid cloning vector derived from pBR322 but improved for easier cloning and screening
Contains bla gene for ampicillin resistance as a selection marker but also contains a multiple cloning site (MCS)
A MCS is a short (50-75 bp) synthetic DNA region containing many unique restriction enzymes so that the sites only occur once in the plasmid which prevents unwanted cutting elsewhere
The MCS region is inserted inside lacZα which enables α-complementation and blue-white screening
lacZ encodes β-galactosidase which is an enzyme that breaks down lactose
β-galactosidase has a α-fragment and a ω-fragment that together form the active form of the enzyme
In this system, the bacterial chromosome contains lacZΔM15 which only makes the ω-fragment so it cannot produce a functional enzyme alone
The pUC19 plasmid carries lacZα which produces the α-fragment
If the plasmid is intact, functional β-galactosidase, a process known as α-complementation
Recombinant clones are then identified using Blue-White screening
The plate contains ampicillin to select for bacteria containing the plasmid as if there is no plasmid the cell will die
It also contains IPTG which is an artificial inducer of the lac operon to turn on lacZ expression
Finally, it also contains X-gal which is a colourless substrate for β-galactosidase, if it is active then X-gal is cleaved and blue pigment forms
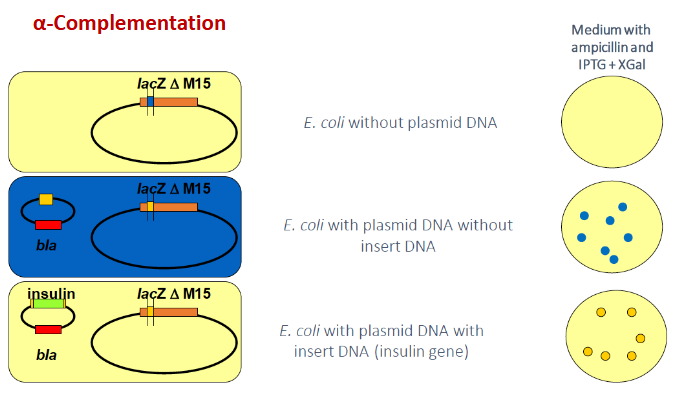
No plasmid → cells lack AmpR and die on the ampicillin plate
Plasmid without insert → lacZα is intact, α-complementation occurs so X-gal is cleaved and colonies turn blue which identifies them as non-recombinant plasmids
Plasmids with insert DNA → foreign DNA inserted into MCS disrupts lacZα so no α-fragment produced and no functional functional β-galactosidase so X-gal is not cleaved and colonies remain white which identifies them as recombinant clones