Matriz extracelular animal y pared celular vegetal
Diversidad de la matriz extracelular (ECM)
La ECM animal adopta formas muy distintas, regulando no solo la forma y las propiedades mecánicas de tejidos y órganos, sino también influyendo en la proliferación, diferenciación y supervivencia celular.
Hueso: Se caracteriza por una matriz extracelular extremadamente rígida y calcificada, compuesta principalmente por fosfato de calcio. Contiene muy pocas células (osteocitos) embebidas, lo que la hace un tejido de soporte principal y reserva de minerales.
Cartílago: Posee una matriz casi íntegra pero notablemente flexible, rica en proteoglucanos (como el agrecán) y fibras de colágeno (principalmente tipo II). Esta composición le confiere resistencia a la compresión y elasticidad.
Tejido conectivo laxo (dermis, alrededor de glándulas y vasos sanguíneos): Presenta una matriz gelatinosa y abundante, con una alta proporción de ácido hialurónico y otras moléculas, que permite el paso de nutrientes, oxígeno y células inmunitarias. Contiene numerosos fibroblastos que secretan y organizan esta matriz.
Epitelios: No solo descansan sobre, sino que secretan activamente una ECM altamente especializada conocida como lámina basal, que les proporciona soporte estructural y señales para mantener su polaridad y diferenciación.
Tres clases universales de moléculas en la ECM que interactúan de forma compleja para formar una red tridimensional dinámica:
Proteínas estructurales: Incluyen los colágenos, que proporcionan resistencia a la tensión y la elastina, que aporta elasticidad y flexibilidad al tejido.
Complejos proteína-polisacárido (proteoglucanos): Son macromoléculas que consisten en una proteína central a la que se unen numerosas cadenas de glicosaminoglucanos (GAGs). Estos complejos son altamente hidrofílicos y generan una fase gel que resiste fuerzas de compresión y facilita la difusión de moléculas.
Glucoproteínas adhesivas: Como la fibronectina y la laminina, que actúan como "pegamento" molecular, conectando las células a los componentes de la ECM y viceversa, facilitando la comunicación bidireccional entre la célula y su entorno.
Colágenos: fuente de resistencia tensil
El colágeno es la familia más abundante de proteínas en la ECM y, de hecho, la proteína más abundante de todo el cuerpo vertebrado, representando aproximadamente el del total proteico corporal.
Son secretados principalmente por fibroblastos en el tejido conectivo, pero también por otras células como osteoblastos (en hueso) y condroblastos (en cartílago).
Poseen dos rasgos moleculares clave que determinan sus propiedades:
Una estructura de triple hélice rígida formada por la superposición helicoidal de tres cadenas polipeptídicas individuales (cadenas ), que giran en sentido derecho. Esta estructura confiere la alta resistencia a la tracción.
Una composición inusual de aminoácidos: son extraordinariamente ricos en glicina (Gly) y en los aminoácidos modificados post-traduccionalmente hidroxiprolina (HyP) e hidroxilisina (HyL). La glicina, debido a su pequeño tamaño (solo un átomo de hidrógeno como cadena lateral), es el único aminoácido que puede ocupar el eje interior estrecho de la triple hélice, siendo esencial para su formación y estabilidad. La hidroxiprolina y la hidroxilisina, formadas por la acción de enzimas hidroxilasas (que requieren vitamina C), son cruciales para la formación de puentes de hidrógeno inter-cadena y la reticulación, lo que aumenta significativamente la estabilidad de la molécula.
Las dimensiones de una molécula de tropocolágeno individual (la unidad monomérica de colágeno) son: una longitud y un diámetro .
El ensamblaje del colágeno es un proceso complejo que ocurre tanto intracelular como extracelularmente (Figura 15-13):
En el Retículo Endoplasmático (R.E.): Tres cadenas de procolágeno (precursores con péptidos en sus extremos N- y C-terminales que impiden el autoensamblaje prematuro) se enrollan para formar una triple hélice, dando lugar al procolágeno. La hidroxilación de Pro y Lys ocurre en este paso.
Secreción y procesamiento extracelular: El procolágeno es secretado al espacio extracelular, donde enzimas específicas llamadas procolágeno-peptidasas cortan los péptidos terminales de procolágeno, transformándolo en colágeno (o tropocolágeno).
Auto-asociación y formación de fibrillas y fibras: Las moléculas de colágeno se auto-asocian espontáneamente en un patrón escalonado y compacto (desfasadas en un cuarto de su longitud), formando fibrillas de colágeno. Estas fibrillas presentan una periodicidad característica de estrías de aproximadamente 67 nm bajo el microscopio electrónico. Múltiples fibrillas se agrupan lateralmente para formar fibras macroscópicas, visibles al microscopio óptico.
Puentes de hidrógeno y reticulación: Los grupos hidroxilo (OH) de la hidroxiprolina y la hidroxilisina forman numerosos puentes de hidrógeno intra- e inter-moleculares, estabilizando la triple hélice y promoviendo la reticulación covalente entre moléculas de colágeno adyacentes, lo que incrementa aún más la estabilidad y resistencia mecánica del tejido.
La resistencia mecánica del colágeno es impresionante: se requiere una carga superior a para romper una sola fibra de colágeno de 1 mm de diámetro.
Existe una gran diversidad génica de colágenos:
Aproximadamente 25 genes distintos codifican para las cadenas , lo que da lugar a al menos 15 tipos de colágeno diferentes (Tabla 15-3), cada uno con patrones de ensamblaje (formadores de fibrillas, formadores de redes, asociados a fibrillas, etc.), distribución tisular y funciones específicas.
El colágeno tipo I es el más abundante en humanos, constituyendo aproximadamente el del colágeno total, y se encuentra en tejidos como piel, hueso, tendones y ligamentos.
Patologías asociadas con el colágeno demuestran su importancia funcional:
Síndrome de Ehlers–Danlos: Es un grupo heterogéneo de trastornos hereditarios del tejido conectivo causados por mutaciones en genes que codifican para colágenos (ej., tipo I, III, V) o en enzimas implicadas en su procesamiento. Se manifiesta con hiperlaxitud articular (articulaciones excesivamente flexibles), piel frágil y elástica, y fragilidad de vasos sanguíneos, lo que puede llevar a aneurismas y hemorragias internas.
Escorbuto: Es una enfermedad nutricional causada por un déficit grave de vitamina C (ácido ascórbico). La vitamina C es un cofactor esencial para las enzimas prolil hidroxilasa y lisil hidroxilasa, que catalizan la hidroxilación de residuos de prolina y lisina en el procolágeno. Sin una hidroxilación adecuada, las cadenas no pueden formar una triple hélice estable, lo que resulta en un colágeno defectuoso y una consecuente fragilidad de los vasos sanguíneos, hemorragias (encías, piel), dolor articular y lenta cicatrización de heridas.
Elastinas: elasticidad & flexibilidad
Las proteínas de elastina son componentes clave de la ECM que confieren elasticidad a los tejidos, predominando en aquellos que requieren estirarse y recuperar su forma repetidamente, como el pulmón, las paredes arteriales, la piel y el intestino.
Son ricas en los aminoácidos glicina (Gly) y prolina (Pro), pero a diferencia del colágeno, carecen de hidroxilisina. Las moléculas de elastina se unen entre sí de forma covalente, formando enlaces cruzados únicos derivados de residuos de lisina, conocidos como desmosina e isodesmosina, que interconectan hasta cuatro cadenas de elastina, creando una red elástica tridimensional.
El funcionamiento de la elastina como un resorte molecular se basa en su estructura y los enlaces cruzados:
Cuando se aplica una tensión externa, las cadenas de elastina, que en ausencia de tensión están en una conformación relajada y desordenada (aleatoria), se estiran y adoptan una conformación más extendida y ordenada.
Al liberar la tensión, las numerosas reticulaciones covalentes actúan como puntos de anclaje que inducen el retorno rápido de las moléculas a su estado relajado y desordenado termodinámicamente más favorable, similar al retroceso de una banda de goma.
El envejecimiento afecta negativamente a las propiedades de la ECM:
El colágeno se vuelve más reticulado y, por lo tanto, más inflexible y quebradizo con la edad, contribuyendo a la rigidez de los tejidos.
La elastina se degrada progresivamente y no se repone eficazmente, lo que lleva a una pérdida de elasticidad y turgencia en la piel (resultando en arrugas) y a una mayor rigidez en las articulaciones y los vasos sanguíneos.
Proteoglucanos y GAGs: fase gel hidratada
Definición: Los proteoglucanos son macromoléculas complejas que consisten en una proteína central a la que se unen covalentemente, y de forma repetitiva, numerosas cadenas de glicosaminoglucanos (GAGs) sulfatadas. A menudo, se asemejan a cepillos de botella por su estructura.
Los GAGs son polisacáridos lineales no ramificados, compuestos por unidades disacáridas repetidas (azúcares amino y ácidos urónicos). Los GAGs más comunes y sus dímeros repetidos característicos incluyen (Figura 15-15c):
Condroitín sulfato: Dímero de (ácido glucurónico y N-acetilgalactosamina sulfatada en la posición 6).
Keratán sulfato: Dímero de (galactosa y N-acetilglucosamina sulfatada en la posición 6).
Hialuronato (o ácido hialurónico): Es el GAG más grande y único que no está sulfatado ni se une covalentemente a una proteína central para formar un proteoglucano individual. Su dímero repetido es (ácido glucurónico y N-acetilglucosamina). Se sintetiza en la membrana plasmática y se libera directamente al espacio extracelular.
La característica distintiva de los GAGs es su alta densidad de cargas negativas (debido a los grupos sulfato y carboxilo en sus azúcares). Estas cargas atraen grandes cantidades de moléculas de agua (formando un gel hidratado) y cationes (como el sodio), creando una presión osmótica interna considerable. Esto permite que la matriz extracelular resista fuerzas de compresión y actúe como un amortiguador, además de facilitar la difusión de nutrientes, metabolitos y pequeñas moléculas de señalización.
El tamaño y la complejidad de los proteoglucanos pueden variar enormemente:
El peso molecular de su núcleo proteico puede oscilar desde hasta .
Pueden tener entre 1 y 200 cadenas de carbohidratos (GAGs), cada una compuesta por aproximadamente 800 azúcares.
El peso molecular total de un proteoglucano puede ir desde hasta , destacando su naturaleza macromolecular y su capacidad de ocupar grandes volúmenes.
En el cartílago, los proteoglucanos, especialmente el agrecán, forman agregados gigantes al unirse al hialuronato a través de proteínas enlace. Esta estructura masiva es fundamental para la resiliencia del cartílago, permitiéndole soportar y recuperar su forma tras cargas de compresión intensas, funcionando como un "amortiguador biológico".
Funciones adicionales de los proteoglucanos:
Actúan como co-receptores en señalización celular, interactuando con factores de crecimiento y citocinas. Por ejemplo, los sindecanes (proteoglucanos transmembrana) con cadenas de heparán sulfato, pueden modular la señalización de factores de crecimiento como el FGF (factor de crecimiento fibroblástico), presentando el factor al receptor específico en la superficie celular y regulando su actividad.
El Hialuronato libre (no asociado a proteína central) tiene funciones importantes por sí solo, actuando como un excelente lubricante en las articulaciones (componente principal del líquido sinovial) y como un medio que facilita la migración celular durante el desarrollo embrionario, la cicatrización de heridas y la remodelación tisular, al proporcionar un espacio hidratado y una vía de baja resistencia para el movimiento celular.
Glucoproteínas adhesivas
Fibronectina (FN)
La fibronectina es una glucoproteína adhesiva de la ECM que se presenta como un dímero grande, con cada subunidad de aproximadamente 2500 aminoácidos, unidas covalentemente por dos puentes disulfuro cerca de su extremo C-terminal.
Su estructura consiste en una serie de dominios modulares (aproximadamente 30) interconectados por segmentos flexibles. Esta modularidad le permite tener múltiples sitios de unión para diversas moléculas:
Colágeno: Puede unirse a varios tipos de colágeno, incluyendo el tipo I, II y IV.
Heparina: Se une a proteoglucanos con cadenas de heparán sulfato.
Fibrina: Interviene en la coagulación sanguínea al unirse a la fibrina, formando parte del coágulo sanguíneo.
Receptores celulares: Contiene secuencias específicas reconocidas por receptores de la superficie celular, como el motivo RGD (Arg-Gly-Asp), que es el sitio de reconocimiento principal para la mayoría de las integrinas (receptores de adhesión celular).
La fibronectina existe en dos formas principales:
Fibronectina soluble (plasma FN): Producida principalmente por los hepatocitos, se encuentra en grandes cantidades en el plasma sanguíneo y otros fluidos corporales. Participa activamente en procesos como la coagulación sanguínea (uniéndose a plaquetas y fibrina), la cicatrización de heridas y el transporte de moléculas.
Fibronectina insoluble: Se ensambla en forma de fibrillas en la matriz extracelular (ECM) por los fibroblastos y otras células, formando una red estructural que guía la organización de otras proteínas de la ECM y proporciona un andamiaje para la adhesión y migración celular.
Funciones cruciales y ejemplos de su implicación biológica:
Migración embrionaria: Desempeña un papel fundamental en el desarrollo, ya que muchas células embrionarias (ej., células de la cresta neural) migran siguiendo caminos ricos en fibrillas de fibronectina, que actúan como guías del desarrollo.
Experimentos de "knock-out" en ratón: La eliminación génica de fibronectina en modelos murinos es letal en etapas tempranas del desarrollo embrionario, demostrando su esencialidad para la formación y el desarrollo normal de estructuras musculares y vasculares (por fallos severos en la morfogénesis del corazón y vasos sanguíneos).
Cáncer: En muchos tipos de tumores malignos, las células cancerosas pierden o producen menos fibronectina de la forma fibrilar. Esta deficiencia contribuye a la pérdida de forma celular, la reducción de la adhesión célula-matriz y un aumento de la movilidad y capacidad invasiva (metástasis). Interesantemente, la adición de fibronectina exógena a estas células tumorales puede, en algunos casos, inducir la recuperación de su morfología normal y la adhesión, sugiriendo un papel supresor de tumores en ciertos contextos.
Laminina
La laminina es la principal glucoproteína adhesiva de la lámina basal, una ECM especializada y fina (aproximadamente 50 nm de grosor) que subyace a todos los epitelios, endotelios y células musculares/adiposas, separándolos del tejido conectivo subyacente.
Es un heterotrímero grande (peso molecular aproximado de ) compuesto por tres cadenas polipeptídicas distintas (, y ) unidas por puentes disulfuro. Su estructura característica es en forma de cruz asimétrica o "T", con un brazo largo central que corresponde a una región de triple hélice y tres brazos cortos. Esta forma permite múltiples interacciones.
La laminina posee múltiples sitios de unión, lo que le permite interactuar con otros componentes de la lámina basal y con las células:
Colágeno IV: El colágeno principal de la lámina basal, al que la laminina se une para formar una red.
Heparina y sulfato de heparán: Se une a proteoglucanos, como el perlecán, que es un componente importante de la lámina basal.
Nidógeno (o entactina): Una glucoproteína que actúa como puente, facilitando la interacción entre laminina y colágeno IV.
Receptores celulares: Posee sitios de reconocimiento para integrinas (como y ) y otros receptores de la superficie celular, mediando la adhesión celular.
La lámina basal es una estructura altamente organizada compuesta por una red compleja de colágeno IV, laminina, proteoglucanos (como perlecán) y nidógeno.
Sus funciones son diversas y críticas:
Soporte estructural: Sirve como un andamiaje para la adhesión, polaridad y organización de las células epiteliales.
Filtración y barrera: Actúa como una barrera selectiva, controlando el paso de moléculas y células. Un ejemplo destacado es la barrera glomerular en el riñón, donde la lámina basal del glomérulo renal filtra el plasma sanguíneo, impidiendo el paso de proteínas grandes y células sanguíneas a la orina.
Guía de migración celular: Proporciona una vía para la migración de ciertas células durante el desarrollo y la reparación tisular.
En cáncer: Las células metastásicas a menudo expresan y secretan MMPs (metaloproteinasas de matriz) en mayor cantidad. Estas son enzimas dependientes de iones metálicos (principalmente zinc) que degradan los componentes de la ECM, incluyendo el colágeno IV y la laminina de la lámina basal. Esta degradación facilita que las células tumorales invadan los tejidos circundantes, penetren en vasos sanguíneos o linfáticos (intravasación) y se diseminen a sitios distantes (metástasis), un paso crucial en la progresión maligna.
Además de la invasión tumoral, las MMPs también desempeñan un papel fisiológico importante en la infiltración leucocitaria durante la respuesta inflamatoria o inmunitaria, permitiendo que las células inmunes atraviesen las barreras tisulares para llegar al sitio de la infección o daño.
Integrinas: receptores mecano-químicos
Las integrinas son la principal familia de receptores transmembrana que median la adhesión de las células a la ECM y la comunicación bidireccional entre la célula y su entorno. Funcionan como heterodímeros no covalentes, compuestos por una subunidad y una subunidad . Existen aproximadamente 18 subunidades y 8 subunidades diferentes en mamíferos.
Las masas moleculares de estas subunidades son: y .
La combinatoria de estas subunidades y genera al menos 24 tipos de integrinas diferentes, cada una con especificidades de unión a ligandos y distribuciones tisulares distintas. Por ejemplo:
La integrina reconoce y se une preferentemente a la fibronectina.
La integrina se une principalmente a la laminina.
La integrina (en combinación con diferentes cadenas ) se expresa casi exclusivamente en leucocitos y es crucial para sus funciones inmunitarias, como la adhesión a las células endoteliales y la extravasación.
Sus dominios extracelulares son responsables de reconocer y unirse a secuencias específicas de la ECM, como el motivo RGD (Arg-Gly-Asp) en fibronectina y otros. Paralelamente, sus dominios citosólicos cortos (aproximadamente 50 aminoácidos) no tienen actividad enzimática intrínseca, pero se unen a diversas proteínas adaptadoras intracelulares, actuando como puentes hacia el citoesqueleto y como plataformas de señalización.
Las integrinas son cruciales para establecer conexiones citosqueléticas que anclan la célula a la ECM y transmiten fuerzas mecánicas:
Adhesiones focales: Son estructuras dinámicas y altamente organizadas que se forman en células migratorias o no epiteliales (como fibroblastos). En estas uniones, las integrinas se unen a la ECM exteriormente y, en su lado citosólico, se conectan a proteínas adaptadoras como talina, vinculina y -actinina, las cuales a su vez se asocian directamente con los filamentos de actina (parte del citoesqueleto). Estas uniones son esenciales para la motilidad celular, la proliferación y la señalización.
Defectos en integrinas que se unen a la laminina (ej. en músculo) o en sus proteínas asociadas pueden llevar a distrofias musculares severas, resaltando el papel mecánico y de señalización crítico de estas conexiones.
Hemidesmosomas: Son uniones de adhesión robustas y estables que se encuentran exclusivamente en las células epiteliales basales, anclándolas firmemente a la lámina basal subyacente. En los hemidesmosomas, una isoforma específica de integrina, la , se une a la laminina en la lámina basal. En la cara citosólica, interactúa con proteínas como la plectina y BPAG (antígeno de pénfigo bulloso), que a su vez ligan directamente los filamentos intermedios (específicamente de queratina en epitelios). A diferencia de las adhesiones focales, los hemidesmosomas están diseñados para una adhesión fuerte y estática, crucial para la integridad estructural del epitelio.
Las integrinas son verdaderos receptores mecano-químicos que participan activamente en la señalización celular bidireccional:
"Inside-out" signaling: Señales intracelulares (ej. activación de GTPasas como Rap1) pueden modular la conformación de las integrinas, alterando su afinidad por el ligando en la ECM y promoviendo su agrupamiento (clustering) en la superficie celular. Esto permite a la célula adaptar su adhesión y migración en respuesta a estímulos internos.
"Outside-in" signaling: La unión de las integrinas a sus ligandos en la ECM y su posterior agrupamiento desencadena la activación de diversas vías de señalización intracelular, incluyendo quinasas como la FAK (quinasa de adhesión focal), ILK (quinasa ligada a integrina), kindlin y paxilina. Estas vías controlan procesos celulares fundamentales como la proliferación, la supervivencia (inhibiendo la apoptosis), la diferenciación y la migración celular. Las integrinas, por lo tanto, no solo anclan la célula, sino que también informan a la célula sobre su entorno mecánico y químico.
Crecimiento dependiente de anclaje: La mayoría de las células normales dependen de la adhesión a una ECM para proliferar y sobrevivir. Si se les priva de esta adhesión (ej., si se cultivan en suspensión sin una superficie de unión), inducen un tipo de muerte celular programada llamado anoikis (apoptosis inducida por la falta de anclaje). Las células cancerosas, sin embargo, a menudo evaden este control, lo que les permite sobrevivir y proliferar en ausencia de anclaje, un rasgo distintivo de la transformación maligna. Esto a menudo se debe a una activación constitutiva de vías de señalización aguas abajo de las integrinas, como la activación de FAK, incluso sin una adhesión adecuada a la ECM.
Costameros y distrofia muscular
El costamero es una compleja zona de adhesión circunferencial en la membrana plasmática de las células musculares estriadas (esqueléticas y cardíacas). Se alinea con los discos Z del sarcómero (unidad contráctil del músculo), actuando como un "anclaje" lateral. Su función principal es conectar las miofibrillas contráctiles (compuestas por actina y miosina) con la membrana plasmática y, a través de ella, con la matriz extracelular (ECM).
Esta conexión se logra a través del complejo distrofina/distroglicano, que es el enlace molecular clave:
La distrofina es una proteína citosólica grande (de aproximadamente <100 nm de longitud) que une los filamentos de F-actina (el citoesqueleto contráctil) en el lado interno de la membrana plasmática.
La distrofina, a su vez, se une a un complejo de proteínas transmembrana conocido como distroglicanos (formado por y distroglicano).
Los distroglicanos, en su dominio extracelular, interactúan directamente con la laminina (específicamente la laminina-211) en la matriz extracelular (lámina basal muscular) que rodea la fibra muscular. Así, el costamero forma un puente mecánico y de señalización desde el interior de la célula muscular hasta el exterior.
Mutaciones en el gen que codifica la distrofina son la causa principal de las distrofias musculares de Duchenne (DMD) y Becker (BMD), dos enfermedades neuromusculares progresivas. DMD es la forma más severa, con una incidencia de aproximadamente varones (es un rasgo ligado al cromosoma X). Estas mutaciones llevan a una proteína distrofina ausente, truncada o disfuncional, lo que interrumpe la conexión entre las miofibrillas y la ECM. Esto resulta en una debilidad muscular progresiva (esquelética y cardíaca), daño muscular crónico, degeneración tisular y, eventualmente, muerte temprana debido a insuficiencia respiratoria o cardiomiopatía.
La función mecánica del costamero y del complejo distrofina/distroglicano es la de un "resorte rígido" o "amortiguador". Evitan el daño inducido por el estrés mecánico y las fuerzas de cizallamiento generadas durante la contracción muscular repetitiva, distribuyendo estas fuerzas de manera uniforme sobre la superficie de la fibra muscular y transmitiéndolas a la ECM. Cuando este sistema falla, las microlesiones se acumulan, llevando a la degeneración de la fibra muscular y su reemplazo por tejido fibroso y adiposo.
Cultivos 3D, Matrigel y bioingeniería (Key Technique)
Problema: Los cultivos celulares tradicionales en placas de plástico (cultivos 2D) no replican adecuadamente la complejidad biofísica y bioquímica de la ECM in vivo. Las células en 2D a menudo pierden su polaridad, diferenciación y comportamiento fisiológico, limitando la relevancia de los resultados de investigación y el desarrollo de terapias.
Solución: El uso de matrices 3D, como el Matrigel, ha revolucionado la investigación celular. El Matrigel es un extracto solubilizado de la lámina basal de un tumor de ratón (sarcoma de Engelbreth-Holm-Swarm, EHS) que contiene una mezcla rica de componentes de la ECM como laminina, colágeno IV, heparán sulfato, proteoglucanos y factores de crecimiento. Puede usarse como una fina capa para imitar una lámina basal 2D o como una matriz 3D para permitir que las células crezcan en un entorno tridimensional.
Ejemplo ilustrativo: Las células epiteliales mamarias no transformadas (línea celular MCF-10A) se utilizan comúnmente para demostrar la importancia del entorno en 3D:
En cultivo 2D: Las células MCF-10A crecen como una simple monocapa "empedrada" sobre el plástico, mostrando una morfología aplanada y una polaridad apical-basal modificada.
En cultivo 3D en Matrigel: Cuando se siembran en un gel de Matrigel, las mismas células MCF-10A son capaces de auto-organizarse y formar quistes epiteliales polarizados que imitan las estructuras glandulares mamarias in vivo. Dentro de estas estructuras, las células establecen un lumen central, adquieren una polaridad apical-basal definida y son capaces de diferenciarse y producir proteínas de leche (en respuesta a señales hormonales), demostrando cómo la ECM tridimensional dirige la morfogénesis y la función.
Importancia: Esta técnica ha sido crucial por varias razones:
Revela el control de forma y diferenciación por la ECM: Ha demostrado de manera contundente cómo la composición y arquitectura de la ECM (incluyendo su rigidez, porosidad y composición bioquímica) influyen profundamente en la forma celular, la diferenciación, la proliferación y el comportamiento maligno.
Aplicaciones en ingeniería tisular y medicina regenerativa: Facilita el desarrollo de tejidos y órganos funcionales in vitro. Se utiliza en la bioingeniería para construir andamiajes y matrices que promueven el crecimiento y la diferenciación de células para la regeneración de diversos tejidos, incluyendo válvulas cardíacas, segmentos de tráquea, injertos de piel y estructuras cartilaginosas como el pabellón auricular. Ofrece un camino prometedor para la reparación y el reemplazo de tejidos dañados.
Pared celular vegetal: analogía y diferencias
La pared celular vegetal es una estructura extracelular rígida y resistente que rodea la membrana plasmática de las células vegetales, hongos, algas y bacterias (aunque con composiciones muy diferentes). En las plantas, a diferencia de la ECM animal, la pared celular proporciona el esqueleto externo primario de la célula y de la planta entera, resiste la elevada presión de turgencia interna de la célula (manteniendo su forma y evitando la lisis osmótica) y actúa como una barrera protectora contra patógenos y el estrés ambiental, así como controlando la expansión celular.
Su composición típica (en peso seco) varía según el tipo de célula y la etapa de desarrollo, pero generalmente incluye:
Celulosa (aproximadamente ): Un polisacárido estructural formado por microfibrillas. Es el componente más abundante.
Hemicelulosas (aproximadamente ): Polisacáridos de cadena ramificada (ej., xiloglucanos, arabinoxilanos) que se unen a las microfibrillas de celulosa y las entrecruzan, aumentando la resistencia tensil de la red.
Pectinas (aproximadamente ): Polisacáridos altamente hidratados y ramificados, con cargas negativas, que forman una matriz gelatinosa que rellena los espacios entre las microfibrillas de celulosa y las hemicelulosas, contribuyendo a la resistencia a la compresión y la porosidad de la pared.
Glucoproteínas como las extensinas (aproximadamente ): Proteínas ricas en hidroxiprolina que pueden reticularse y fortalecer la pared, especialmente durante el desarrollo.
Lignina: Un polímero complejo e hidrofóbico que se deposita en la pared celular de los tejidos leñosos (madera) para aportar una rigidez y resistencia mecánica adicional, crucial para el soporte de plantas grandes.
La arquitectura de la pared celular vegetal (Figura 15-22) es comparable a un hormigón armado, donde las microfibrillas de celulosa actúan como las varillas de acero (el elemento de resistencia a la tensión) y las hemicelulosas y extensinas forman una red que las interconecta firmemente. Toda esta estructura está incrustada en una matriz de pectina gelatinosa, que proporciona resistencia a la compresión y actúa como un medio de difusión. Las microfibrillas de celulosa están compuestas por cadenas lineales de glucosa unidas por enlaces glucosídicos , que se agrupan en microfibrillas (de unos de diámetro) para formar macrofibrillas aún mayores.
Comparación entre ECM animal y pared vegetal (Tabla 15-2): Aunque funcionalmente análogas en su papel de soporte estructural y mediación del entorno celular, sus composiciones moleculares son fundamentalmente diferentes:
Animal ECM: Los principales elementos son el colágeno (resistencia a la tensión) y la elastina (elasticidad), los proteoglucanos (resistencia a la compresión y fase gel), y las fibronectina/laminina (adhesión y organización).
Pared celular vegetal: Los elementos análogos son la celulosa (resistencia a la tensión), las hemicelulosas/extensinas (entrelazado y resistencia), y las pectinas (fase gel y resistencia a la compresión).
Conexiones, implicaciones filosóficas y éticas
La profunda influencia de la ECM (y en plantas, de la pared celular) en el destino celular (decidiendo si una célula prolifera, se diferencia, migra o muere) nos recuerda que la identidad y el comportamiento celular no residen únicamente en la información genética contenida en el genoma, sino que están inextricablemente ligadas al contexto ambiental tridimensional en el que la célula se encuentra. Esto subraya la importancia de la epigenética y las interacciones célula-entorno en la biología.
La degradación excesiva de la ECM, particularmente por MMPs en patologías como el cáncer, plantea importantes dilemas terapéuticos. Mientras que inhibir la actividad de las MMPs podría frenar la metástasis tumoral (al evitar la invasión de la ECM), estas enzimas también cumplen funciones fisiológicas esenciales en el cuerpo (como la remodelación tisular, cicatrización de heridas, angiogénesis e inmunidad). Por lo tanto, el diseño de fármacos para inhibir selectivamente la actividad de MMPs patológicas sin afectar negativamente las funciones fisiológicas necesarias sigue siendo un desafío significativo en la oncología y otras áreas de la medicina.
El avance de la ingeniería tisular y la creación de órganos "de laboratorio" (como páncreas, hígados o riñones parciales) a partir de células y matrices bioingenierizadas abre complejos debates bioéticos. Estos incluyen cuestiones sobre la definición de vida, la modificación de organismos, la equidad en el acceso a estas tecnologías médicas avanzadas y costosas, y la posibilidad de su uso no terapéutico, lo que requiere un marco ético y regulatorio cuidadoso.
Concept Checks & preguntas de examen
¿Cuáles son las ventajas de anclar las uniones adhesivas (como adhesiones focales y hemidesmosomas) al citoesqueleto?
Respuesta: Permite la transmisión y reparto de fuerzas mecánicas desde el exterior de la célula a su interior, asegurando la integridad mecánica del tejido. Facilita la señalización bidireccional entre la célula y la ECM, influyendo en la expresión génica y el comportamiento celular. Contribuye a la organización polarizada de la célula, esencial para la función de tejidos como los epitelios.
¿Por qué una misma célula, bajo condiciones fisiológicas, generalmente no presenta simultáneamente hemidesmosomas y adhesiones focales?
Respuesta: Porque estas estructuras reflejan y refuerzan estados celulares y destinos distintos. Los hemidesmosomas son característicos de células epiteliales estáticas, ancladas firmemente a la lámina basal, que buscan estabilidad y polaridad basal. Las adhesiones focales son típicas de células migradoras (ej., fibroblastos) o no epiteliales que requieren una adhesión más dinámica y la capacidad de remodelar el citoesqueleto de actina para el movimiento. La célula adopta uno u otro fenotipo modulando la expresión de isoformas específicas de integrina y la organización de su citoesqueleto en respuesta a su entorno.
Relacione el déficit de vitamina C con la estabilidad del colágeno y la patología que causa.
Respuesta: La vitamina C es un cofactor esencial para las enzimas prolil hidroxilasa y lisil hidroxilasa, que hidroxilan residuos de prolina y lisina en el procolágeno. La hidroxiprolina y la hidroxilisina son cruciales para la formación de puentes de hidrógeno que estabilizan la triple hélice del colágeno y para la reticulación de las fibrillas. Un déficit de vitamina C (escorbuto) lleva a la producción de colágeno defectuoso y menos estable, lo que resulta en la fragilidad de los vasos sanguíneos, hemorragias, mala cicatrización y debilidad del tejido conectivo en general.
Explique cómo las integrinas vinculan las señales mecánicas de la ECM con las vías de señalización de tirosina quinasas intracelulares.
Respuesta: Las integrinas no poseen actividad quinasa intrínseca, pero actúan como andamios. Cuando se unen a ligandos de la ECM y se agrupan (clustering), reclutan y activan quinasas citosólicas como la FAK (quinasa de adhesión focal) y la ILK (quinasa ligada a integrina). Estas quinasas, entre otras, fosforilan residuos de tirosina en diversas proteínas adaptadoras y efectoras (como paxilina, talina, vinculina), desencadenando cascadas de señalización que afectan la organización del citoesqueleto, la expresión génica, la proliferación celular, la supervivencia y la migración. De esta manera, las fuerzas mecánicas ejercidas sobre la ECM se traducen en respuestas bioquímicas celulares. Este proceso es vital para el crecimiento dependiente de anclaje, donde la falta de adhesión suprime estas vías y puede inducir apoptosis (anoikis).
Conceptos Clave:
ECM (Matriz Extracelular): Es el "andamio" y "pegamento" que rodea las células animales, dando forma a los tejidos y controlando cómo las células se comportan, crecen y sobreviven.
Definición fácil: Es la red de fuera de la célula que le da soporte y le dice qué hacer.
Colágenos: Son las proteínas más abundantes en el cuerpo; actúan como "cuerdas" que dan resistencia y fuerza a los tejidos.
Definición fácil: Las proteínas que nos hacen fuertes y resistentes.
Elastinas: Proteínas que permiten que los tejidos se estiren y vuelvan a su forma original, como un "resorte".
Definición fácil: Las proteínas que nos dan elasticidad, como en la piel.
Proteoglucanos: Son moléculas grandes formadas por una proteína central unida a muchos azúcares (GAGs). Absorben mucha agua y hacen que la matriz sea un "gel" resistente a la compresión.
Definición fácil: Son esponjas en la matriz que retienen agua y amortiguan.
GAGs (Glicosaminoglucanos): Son cadenas de azúcares con muchas cargas negativas, hacen que los proteoglucanos atraigan mucha agua.
Definición fácil: Azúcares que hacen que la matriz sea un gel espeso.
Fibronectina (FN): Una glucoproteína que actúa como "pegamento" conectando las células con el colágeno y otros componentes de la matriz.
Definición fácil: El "cemento" que une las células a su entorno.
Laminina: La "glucoproteína principal" de la lámina basal (una ECM especial) que ayuda a las células epiteliales a adherirse y organizarse.
Definición fácil: Clave para la base de las células que recubren superficies.
Integrinas: Son los "receptores" en la superficie de la célula que le permiten unirse a la ECM y recibir señales de su entorno.
Definición fácil: Los ojos y manos de la célula para "sentir" y agarrarse a su exterior.
Anoikis: Un tipo de "suicidio celular" que ocurre cuando las células normales no pueden anclarse a la ECM; las células cancerosas a menudo lo evaden.
Definición fácil: La autodestrucción de las células si no se pueden agarrar a algo.
Costamero: Es una zona especial en las células musculares que conecta las estructuras contráctiles del interior con la ECM de fuera, como un "ancla" lateral.
Definición fácil: El "gancho" que une el músculo por dentro con lo de fuera.
Abreviaciones con Nombre Completo:
ECM: Matriz Extracelular
FN: Fibronectina
RGD: Arg-Gly-Asp (un "motivo" o secuencia específica de aminoácidos: Arginina-Glicina-Ácido Aspártico)
DMD: Distrofia Muscular de Duchenne
BMD: Distrofia Muscular de Becker
MMPs: Metaloproteinasas de Matriz (enzimas que degradan la ECM)
R.E.: Retículo Endoplasmático (un orgánulo celular)
FAK: Quinasa de Adhesión Focal
ILK: Quinasa Ligada a Integrina
BPAG: Antígeno de Pénfigo Bulloso
GlcUA–GalNAc_6S: Ácido glucurónico y N-acetilgalactosamina sulfatada en la posición 6 (Unidad repetida del condroitín sulfato)
Gal–GlcNAc_6S: Galactosa y N-acetilglucosamina sulfatada en la posición 6 (Unidad repetida del keratán sulfato)
GlcUA–GlcNAc: Ácido glucurónico y N-acetilglucosamina (Unidad repetida del hialuronato)
Notas de Clase:
MEC: es una red 3d compleja de macromoleculas organizadas mas alla de la superficie de las mismas celulas que rodean y la sintetizan, o sea es de produccion local.
Las celulas interactuan quimica y mecanicamente con la MEC y ayudan a orientar y organizarla.
Fibrosis → pensar en la MEC degradandose, cicatrizandose y ya no siendo la misma.
MEC se encuentra solo en organismos multicelulares (en plantas se llama pared celular).
Tejido: Agrupacion de celulas, con funciones especificas colectivas que originan del mismo tejido embrionario.
Componentes del tejido → MEC y celulas.
Funciones de la MEC:
Soporte y anclaje para las celulas.
Influye directamente en la estructura, propiedades mecanicas y funcion de los tejids.
Coordina funciones celulares activando vias de señalizacion intracelulares.
Es el reservorio de moleculas señalizadoras: secuestra y modula, regula la tasa de difusion y la concentración de moleculas señalizadoras - (se pueden crear gradientes moleculares).
Hay muchos tipos de MEC:
Hueso
Cartilago
Tejido conectivo > dermis
MEC liquida = plasma de la sangre, tejido conectivo liquido.
MEC 3 componentes moleculares:
Proteinas estructurales
Proteoglucanos
Glicoproteinas adhesivas → lamininas y fibrolectinas
Razon por variedades de la MEC:
Tipo de Proteinas estructurales
Tipo de proteoglucanos
Relacion de proteinas estructurales y proteoglucanos
Tipo y cantidad de las glicoproteinas adhesivas
Scurvy / Escorbuto: enfermedad por falta de Vitamina C.
Hemorragia gingival → falta de colageno, causa hemorragias facilmente.
Hay 42 genes que codifican para la triple helice del colageno
Solo se han hallado 28-29 tipos de colageno
Las tres cadenas de la triple helice son cadenas alfa (1,2,3), pueden ser homotrimericas o hetero trimericas. (mismas 3 cadenas alfas o 2 iguales y 1 diferente)
Homotrimericas serian tres helices alfa 1
Heterotrimericas serian dos helices alfa 1 y la tercera alfa 2
Gly-X-Y, donde usualmente X es prolina y Y es hidroxiprolina o hidroxilisina.
Glycina (Gly) es mas importante y abundante porque como es mas corta, permite que se compacte y se junten mas las helices.
1. Cadena precursora ) 3 cadenas se ensamblan → 2. procolageno (triple helice con extremos sueltos) ) procolageno peptidasa → 3. tropocolageno / molecula de colageno ) ensamblaje → fibrilas de colageno VER PAGINA 3
Los puentes de disulfuro en las N terminal son solo entre las cadenas mismas, en la C terminal se mira puentes de disulfuro entre las cadenas, esto es para que se mantengan alineadas para ser enroscadas, y luego de ser enroscadas son cortadas. En el momento que se enroscan las tres cadenas, formando una triple helice, se forma el tropocolageno.
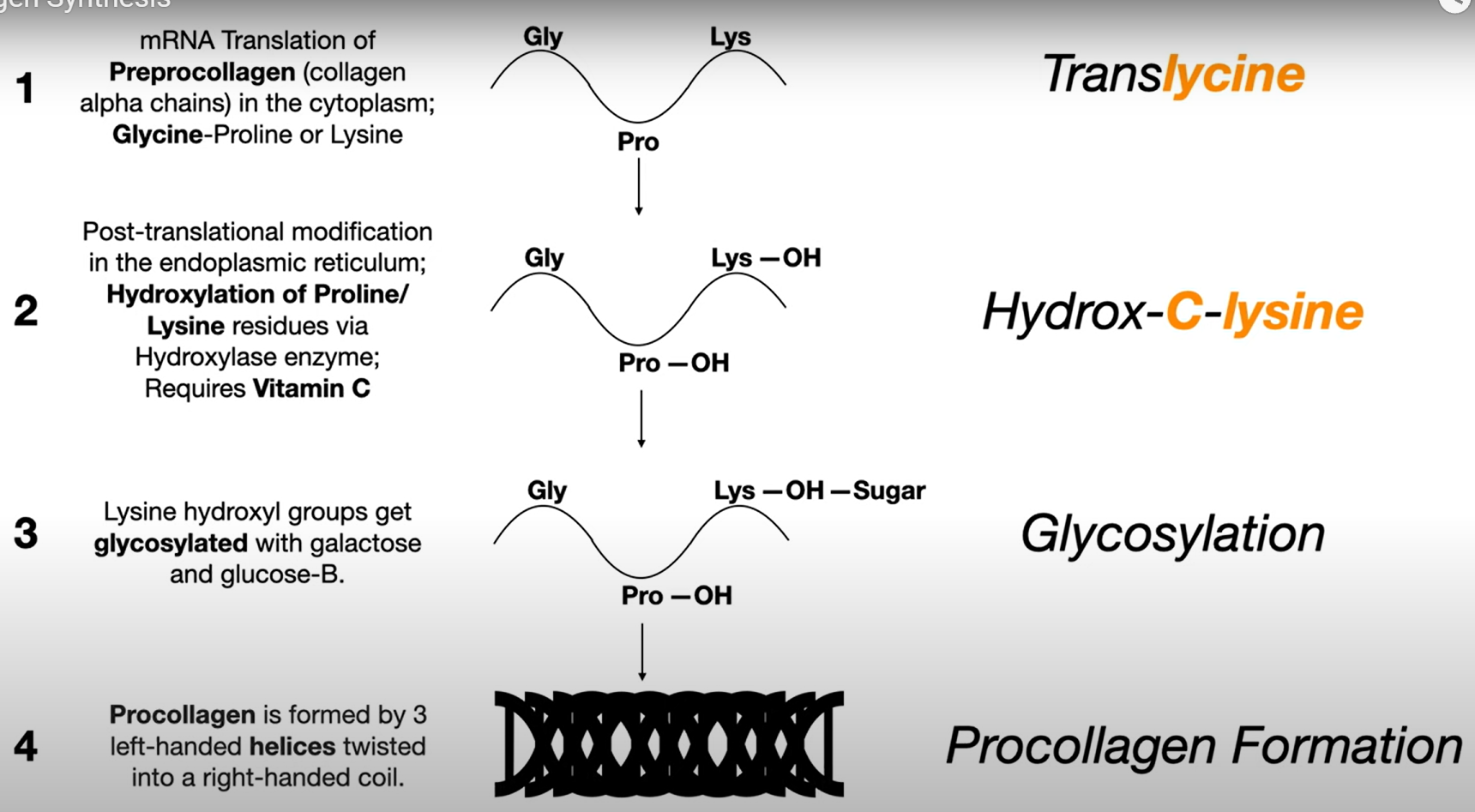
Procolageno peptidasa corta los extremos N y C terminal, pero son diferentes tipos los que corta cada uno.
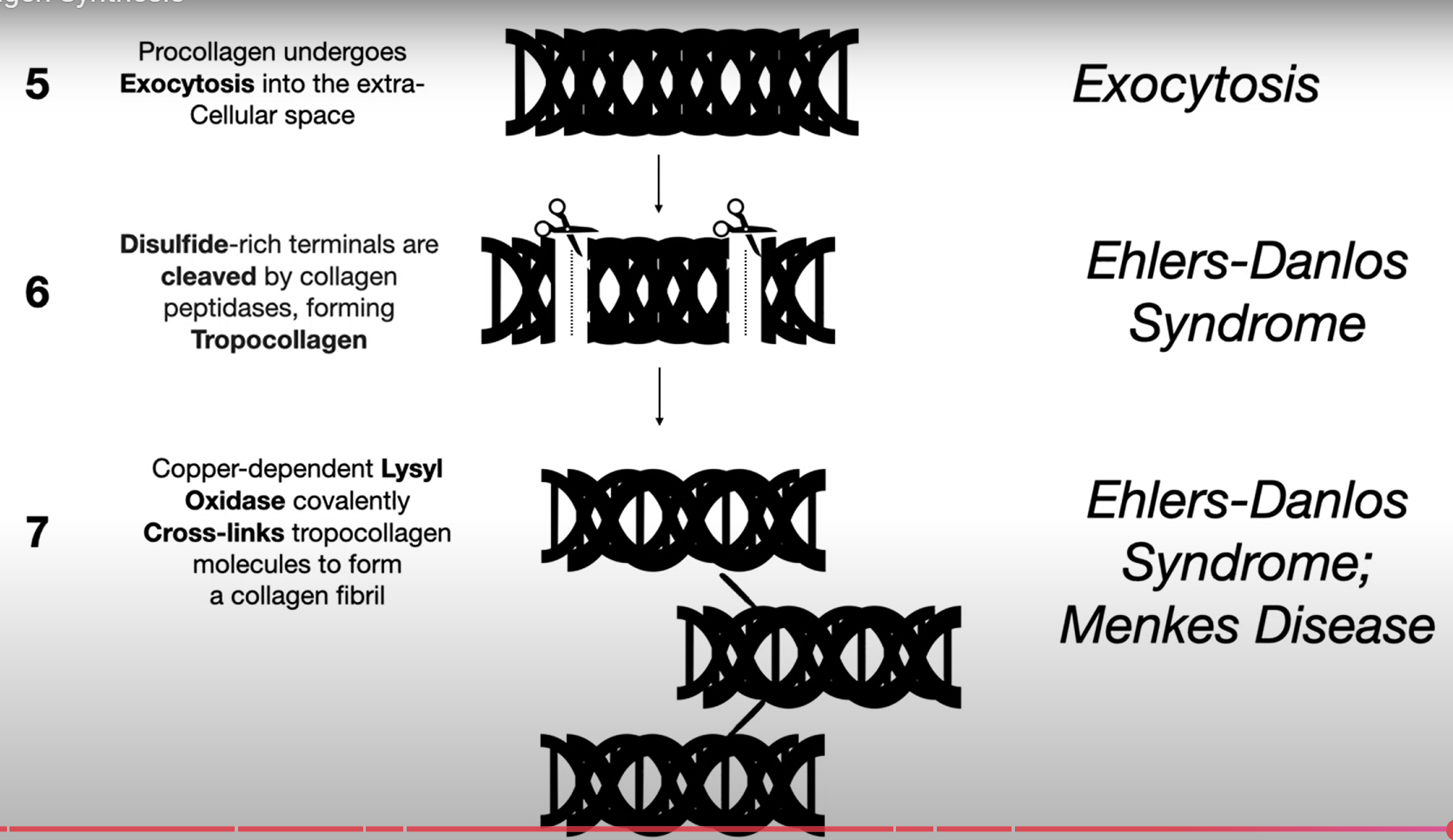
Los tropocolagenos que se unen entre si forman fibrillas, y las fibrillas se unen para formar fibras. Los tropocolagenos se unen entre si a traves de enlaces covalentes, y estos son unidos covalentemente gracias a la lisil-oxidasa, que cataliza la oxidación de residuos de lisina (e hidroxilosina) y también es crucial para la maduración de las fibras de colágeno, asegurando así la estabilidad y resistencia de la matriz extracelular.
Los enlaces covalentes son lisina-lisina, lisina-hidroxilisina.
Los puentes de hidrogeno de todas las hidroxilaciones son lo que le da la estabilidad al tropocolagen (la triple helice).
La lisil oxidasa necesita el cobre (Cu) como cofactor.
Menkes disease es causado por porque la proteina que transporta el Cu a la lisil-oxidasa está mutada, lo que resulta en una deficiencia en la formación de colágeno y daño a los tejidos conectivos. Problemas de estabilidad de fibrillas.
Peptidil lisil hidroxilasa es la enzima responsable de la hidroxilación de los residuos de lisina en el colágeno, lo que es crucial para su estabilidad y funcionalidad.
Analogia
RER = huesped → agua + limon → dulce → trenza → se va
En el RER = traduccion y translasion, → hidroxilasiones (agregar OH) a lysina y prolina → glucosilacion → etc
Colagenos mas abundantes en el cuerpo: tipo 1, 2 y 3
Colageno tipo 1 es de hueso y piel
Colageno tipo 2 es de cartilago
Colageno tipo 3 es de tejido liso como lo que se encuentra en el higado… en organos
Sindrome de Ehlers-Danlos
90% de ehlers-danlos es el de hED (Erhlers-Danlos Hipermovil), que afecta la producción de colágeno, provocando una hipermovilidad articular y una fragilidad en la piel. Además, este síndrome puede llevar a complicaciones relacionadas con la ruptura de vasos sanguíneos, lo que aumenta el riesgo de hematomas y otras lesiones internas. Formacion de estrillas (fragilidad), muchos moretones, problemas de valvulas cardiacas, hernias de la pared abdominal…
7CED (Ehlers-Danlos tipo 7C) → fragilidad de la piel severa. Ocurre por mutacion de procolageno peptidasa N-terminal.
Fibras elasticas: halladas en pulmon, piel, tubo digestivo, vasos sanuineos grandes, arterias, venas, y cartilago, y etc.
propiedades
extensibilidad - 5xs
fxs
localizacion
estructura = diferentes a las estrucutras del colageno.
Compuesto de elastinas y microfibrillas.
No es lo mismo lastina que fibra elastica.
Elastina: rica en prolina, glicina, lisina y alanina. No poseen hidroxilisina o hidroxiprolina (tal vez un poco de hidroxiprolina).
Precursor de elastina: tropoelastica, la tropoelastina no se llama elastina hasta que haya sido entrecruzadas por enlaces covalentes entre residuos de lisinas (por las lisil oxidasas). El entrecruzamiento permite la elasticidad.
Elastina puede contener aa inusuales como desmosina y allisina (lisina modificada), desmosina siendo importante para las uniones covalentes de lisinas y allisinas.
La microfibrilla se halla antes de la elastina.
La elastina se deposita entre las microfibrillas.
La fibrilina-1 es la que codificada por el gen FBN1
Mutacion del Gen FBN1 da Sindrome de Marfan
↨
Sindrome de Marfan : enfermedad de tejido conectivo heredada, autosomico dominante, 75% de los casos es por esto y el otro 25% es causado por mutacion de novo (aparicion espontanea). Lo que mas afecta el sindrome de Marfan es la piel, pulmon, corazon, aorta y el esqueleto. Concecuencias mas peligrosas de este síndrome incluyen complicaciones cardiovasculares como la dilatación de la aorta, que puede provocar desgarros o rupturas, así como problemas ortopédicos como la escoliosis y la luxación del cristalino en los ojos. Los pacientes con esta enfermedad normalmente no presentan sintomas, por eso la ruptura de la aorta tiene 90% de fatalidad, con no puede ser detectado y murene rapidamente en este caso. Los pacientes con Sindrome de Marfan tienen extremidades largas y delgadas, así como una altura superior a la media, lo que contribuye a un aspecto físico característico. Tambien tienen la boca pequeña y menton pequeño.
Relacon entre TGFB (beta) y la Firbrilina-1: TGFB es un factor de crecimiento (transformante beta), la fibrilina ayuda a modular a este factor de crecimiento, por eso con la falta de la firbirlina-1 se produce una sobreexpresión de TGFB, lo que genera un desequilibrio en la matriz extracelular y contribuye al deterioro del tejido conectivo.
Walker-Murdoch = pulgar puede tocar al meñique al wrap around del punto mas gordo de la muñeca.
Steinberg = pulgar sobresalido al hacer puño.
Sindrome de Marfan → ruptura de la aorta: causada por la dilatacion de la aorta y porque se crea un lumen falso en una incision.
Losartan es un bloqueador de la angiotensina 2, es un antipertensivo. EL losartan tambien puede ayudar con la regulacion del TGFB.
Proteoglucanos:
Son glucoproteinas secretadas que poseen al menos 1 GAG unido covalentemente a la proteina central. 95% de su peso total es carbohidrato. Es una glicoproteina con carbohidratos.
No todos las glicoproteinas son proteoglucanos, ya que algunas pueden no contener GAGs o tener una estructura diferente que no cumple con los criterios necesarios para clasificarse como tales, pero todos los proteoglucanos son glicoproteinas. Varian los proteoglucanos en largo de los GAGs y localizacion de la proteina central.
Ejemplos de proteoglucanos: Decorin, Aggrecan, Sybdecan, Glypican. Hallados como proteinas transmembranales y ancladas a GPI.
Propiedades mecanicas: atraen agua las GAGs con cargas negativas. Forma una red altamente hidratada, gelatinosa donde se hallan embebidos los col y f. elasticas.
permiten resistir fuerzas compresivas (como globo lleo de agua)
presion de turgencia (elasticidad o fuerza que tiene la piel hacia fuera por el tejido)
Una forma de detectar deshidratacion (minimo tipo 2), en personas y bebes, es pelliscando la panza del bebe o la mano de una persona y ver si se queda tieza y parada la piel pelliscada, eso es signo de deshidratacion por aucencia de turgencia.
Funciones de los proteoglucanos: union directa a colagenos, union de la MEC a la celula, y rol importante en la señalizacion.
Modular señalizacion: un factor de crecimiento de fibroblastos (FGF) tiene conformacion doble que no es reconocida por los receprotres de FGF, y lo que pasa es que con la ayuda de un proteoglucano, el proteoglucano le cambia la conformacion al FGF y asi es cuando es detectado por el receptor FGF. Modulacion de señalizacion → proteoglucanos.
Proteina central de los proteoglucanos:
Pensar en una pluma de pajaro, el palo en si es la prot central y los GAGs son los strands de plumas.
GAG: polisacaridos largos lineales con unidades de disacaridos especificos que se repiten, se nombran segun la unidad repitente. Una siempre es un amino azucar, N-acetilglucosamina o N-acetilgalactosamina y una de estas dos + un azucar o azucar acida (esto forma un disacarido especifico, son los que se repiten.)
Disacarido de GAG: ejemplo → N-acetilglucosamina + acido glucoronico
Se miraria asi: N-acetilglucosamina + acido glucoronico + N-acetilglucosamina + acido glucoronico + N-acetilglucosamina + acido glucoronico …. asi va
Unico GAG no sulfatado: Hyaluronato.
Los axucares, azucares acidos o amino azucares usualmente poseen grupos sulfatos, en excepcion a hyaluronato.
GAG Hialuronato:
No es sintetizado por el sistema endomembranoso, tiene su propia fabrica de sintesis en la membrana plasmatica.
Forma una gel viscosa aun en bajas concentraciones. Es tan grande que puede existir solo y se le unen otros proteoglucanos. Tiene muchas cargas negativas y puede almacenar bastante agua, y es un exelente lubricante para las articulaciones, en el liquido sinovial.
Agrecano → agregados de agrecano (hallados en el cartilago)
Cartilago: provee alta turgencia (presion de turgencia)
Es tan grande que a el es el que se le unen los proteoglucanos.
Union covalente de GAGs a la Proteina central
Enlace de tetrasacaridos es lo que une el GAG a la Proteina
Glucoproteinas Adhesivas:
actuan como mediadores entre la MEC y celula.
fibronectina y la laminina son las mas comunes de estas.
tienen multiples dominios que reconocen componentes de la MEC y receptores de la superficie ceular.
las lamininas actuan como puente y se miran somo una cruz, anclaje fuerte de a celula a la MEC
Insolubles son fibrillas y se hallan en la MEC
Las solubles se hallan en liquidos corporales como la sangre
Intermedias se unen debilmente a las membranas plasmatica
Fibronectina:
estructura: al menos 27 isoformas a partir de 1 gen, son tejidos especificos, parecen como nunchucks.
La fibronectina es una glucoproteína adhesiva de la ECM que se presenta como un dímero grande, crucial para la adhesión celular y la organización de la matriz.
Su estructura modular (con ≈30 dominios) le permite unirse a diversas moléculas como el colágeno (tipo I, II, IV), heparina, fibrina, y receptores celulares (principalmente integrinas a través del motivo RGD).
Existe en dos formas:
Soluble (plasma FN): Se encuentra en plasma y fluidos, participando en la coagulación sanguínea y la cicatrización de heridas.
Insoluble: Se ensambla en fibrillas en la ECM, formando una red estructural que guía la adhesión y migración celular.
Sus funciones son cruciales:
Fundamental en la migración embrionaria (ej. células de la cresta neural).
Su ausencia (estudios de knock-out en ratón) es letal en desarrollo embrionario debido a fallos en la morfogénesis.
En el cáncer, la pérdida de fibronectina fibrilar contribuye a la pérdida de adhesión y aumento de la invasividad (metástasis), mientras que su adición puede, en algunos casos, restaurar la morfología y adhesión celular normal.
puede unirse a colageno, heparina, etc. Se une a componentes de la MEC como estructuras fibrosas o proteoglicanos, contribuyendo a la estabilidad y funcionalidad del entorno celular.
Funciones: puente entre la MEC y las celulas. Ayuda a organizar a la MEC.
Movimiento celular → permite guiar la migracion celular, heridas y embriogenesis.
Rol en coagulacion, solubles.
FN y cancer: Sin la fibronectina, la celula se desprende de la MEC y cambia la forma y viaja. Esto seria la metastasis del cancer.
Lamininas
tiene 3 cadenas alfa beta y gama, hay 5 cadenas alfa diferentes que son codificadas por 4 cadenas de beta diferentes y 3 cadenas de gamma que codifican estas de beta. Gamma codifica a beta → beta codifica a alfa. 3 × 4 × 5 = 60 diferentes combinaciones (isoformas).
Son capases de autoensamblarse y pueden formar mallas de lamininas, una sola laminina parece como la calavera de un paraguas. Los dominios LG son lo que parece como la u del handle del paraguas. Los dominios LG unen recptores celular (integrinas, sindecanos (tipo de proteoglucano transmembranal, receptor), glicolipidos sulfatados, distroglicanos.
Lamina Basal: es como una barrera para mantener el orden.
Matriz extracelular especializada, localizada en las membranas basales de los epitelios, y encontrado alrededor de celulas adiposas, de schwan, y de tendones y musculos. Grosor es de 50 nanometros, pero en los riñones es de 400 nanometros.
Varian en funcion y composicion segun su localizacion, pero todas poseen:
4 Componentes de la lamina basal:
laminina
colageno tipo 4
periecano
nidogeo o entactina (glicoproteina adhesiva)
Funciones:
soporte estructural
separacion
organizacion de epitelios
conexion mecanica
barrera permeable para solutos y para las celulas.
migracion'
regeneracion de los tejidos luego de sufrir daño (cicatrizacion)
provee un andamio para que las nuevas celulas migren para mantener arquitectura original del tejido
Rol con el cancer → celulas cancerigenas sintetizan mas proteinas de MEC y mas receptores de superficie para que se puedan adherir las celulas a la lamina basal, pero se adhieren tan fuerte a la lamina basal que they break through y llegan a los vasos sanguineos y llega a otros tejidos, permitiendo la metastasis y la propagación del cáncer a diferentes partes del cuerpo.
Lamina basal no es el mismo que membrana basal. Lamina basal tiene la lamina reticular y la basal. Porque el componente de la membrana basal es la lamina basal y la lamina reticular.
Integrinas
Son una familia de receptores de superficie celular, y tambien son moleculas de adhesion celular. Se le unen al citoesqueleto por dentro, aunque indirectamente, y a su vez se une a componentes de la MEC, conecta el interior de la celula con el exterior y esto puede desencadenar vias de señalizacion.
varian en especificidad y tamaño
Funciones:
unir celular con MEC (motivo RGD y otros)
unir celula con otra celula (s) para reptar/migrar
movimiento celular
señalizacion celular
Estructura:
interacciones no covalentes las que unen las unidades Alfa con Beta
dominio citosolico y transmembranal (subu Beta que se unen proteinas adaptadoras que reclutan al citoesqueleto = dominio citosolico) (dominio intracelular tiene sitios re reconocimiento o union para ligandos intracelulares
Ligando se tiene que unir a ambas subunidades aunque la subu Alfa es la que lleva la especificidad.
son calcio dependientes y magnesio dependientes = concentraciones de calcio y mg influyen sobre la afinidad y especificidad de las integrinas.
hay 8 diferentes tipos de subunidades beta y 18 de unidades alfa (18 × 8 = 144 posibles isoformas, pero solo 24 se han visto en los humanos)
Sitios de reconocimiento para los motivos RGD y otros motivos.
La integrina se une a microfilamentos y filamentos intermedios en la subunidad b en intracelular.
Mutacion en las subunidades beta 1, significan muerte temprana del embrion, no hay implantacion. Glanzman disease es una coagulopatia que es causada por la mutacion de la subunidad beta 7 o por ahi.
Integrinas y citoesqueleto
celulas que migran y las NO epiteliales se unen a la MEC por medio de adhesiones focales
son agrupaciones de integrinas que interacuan con el citoesqueleto y permiten la adherencia de las células a la matriz extracelular, facilitando así procesos como la migración celular y la señalización.
Tiene 2 estados: activos e inactivos; en el estado activo, las integrinas están unidas a ligandos en la matriz extracelular, lo que promueve cambios en la estructura del citoesqueleto y favorece la migración celular, mientras que en el estado inactivo, su capacidad de unión se ve limitada, afectando así la respuesta celular a su entorno. Apagada se ve dormida y como de pies cruzados, activado se mira parado y se desliga la subunidad beta para atraer.
Tiene señalizacion outside-in y inside-out. Además, la señalización outside-in se refiere a cómo las interacciones de las integrinas con la MEC pueden desencadenar respuestas dentro de la célula, mientras que la señalización inside-out permite que señales internas modifiquen la afinidad de las integrinas por sus ligandos en la MEC.
Si se activa adentro se separa por fuera, si se activa afuera, se separa por dentro.
Talina compite con la subunidad Alfa para unirse a la subunidad beta, si el alfa se une a la subunidad beta la integrina esta inactivado, si la talina se une a la beta entonces se activa y suelta la alfa a la beta.
Son muy versatiles la integrinas.
Vocabulario
enlaces cruzados
🔹 Son enlaces covalentes resultado de la enzima lisil oxidasa que permiten a la tropoelastina convertirse en fibras elásticas.
vitamina c
🔹 Es necesaria para las hidroxilaciones de las cadenas alfa del colágeno.
colágeno
🔹 Es la proteína más abundante de la matriz extracelular (MEC).
elastina
🔹 Uno de los componentes principales de las fibras elásticas y también una proteína de la MEC que no posee hidroxilisina ni hidroxiprolina.
colágeno maduro
🔹 Es la estructura del colágeno que se forma después del tropocolágeno.
triple hélice
🔹 Es la estructura única característica del colágeno.
soporte
🔹 Es lo que la MEC provee a las células.
matriz extracelular
🔹 Red tridimensional que rodea a las células y les da soporte y estructura a los tejidos.
fibronectinas
🔹 Son glucoproteínas de la MEC que funcionan como moléculas adhesivas.
dinámica
🔹 Palabra que describe que la producción de la MEC es constante y en cambio continuo.
eucariotes
🔹 Tipo de seres en los que se encuentra la matriz extracelular.
procolágeno
🔹 Es la forma de colágeno con extremos no helicoidales antes de salir de la célula.
glicina
🔹 Aminoácido que aparece cada tres posiciones en las cadenas alfa del colágeno.
La MEC (Matriz Extracelular) es una red 3D dinámica de macromoléculas que rodea y es sintetizada por las células. Provee soporte y anclaje, influye en propiedades mecánicas y coordina funciones celulares a través de señalización.
Diversidad y Componentes de la MEC
La MEC adopta formas muy distintas (ej. Hueso rígida y calcificada; Cartílago flexible, rico en proteoglucanos; Tejido conectivo laxo gelatinosa; Epitelios con lámina basal).
Sus 3 clases de moléculas son:
Proteínas estructurales: Colágenos (resistencia a la tensión) y Elastina (elasticidad/flexibilidad).
Complejos proteína-polisacárido: Proteoglucanos (proteína central con GAGs). Formanción de fase gel hidratada que resiste compresión.
Glucoproteínas adhesivas: Fibronectina (FN) y Laminina (conectan células a la ECM).
Colágenos: Resistencia Tensil
Proteína más abundante ( cuerpo). Secretados por fibroblastos.
Estructura clave: triple hélice rígida de 3 cadenas (homo o heterotriméricas). Ricas en glicina (Gly), hidroxiprolina (HyP) e hidroxilisina (HyL).
Proceso de Ensamblaje (Figura 15-13):
Retículo Endoplasmático (R.E.): 3 cadenas de procolágeno (precursor) se enrollan en triple hélice. Hidroxilación de Pro y Lys (requiere vitamina C).
Secreción: Procolágeno va al exterior.
Procesamiento extracelular: Procolágeno-peptidasas cortan extremos, formando colágeno (tropocolágeno).
Auto-asociación: Moléculas de colágeno se agrupan en fibrillas (en patrón escalonado), luego en fibras macroscópicas. Enlaces covalentes cruzados de lisina por lisil-oxidasa (cofactor Cu) dan estabilidad.
Integridad: Puentes de hidrógeno de HyL/HyP estabilizan la triple hélice.
Patologías:
Escorbuto: Deficiencia de vitamina C → colágeno defectuoso, fragilidad vascular, hemorragias.
Síndrome de Ehlers–Danlos: Mutaciones genes de colágeno/enzimas. Hiperlaxitud articular, piel frágil. (Ej., 7CED por mutación procolágeno peptidasa N-terminal).
Enfermedad de Menkes: Deficiencia de Cu afecta lisil-oxidasa, inestabilidad de fibrillas.
Elastinas: Elasticidad y Flexibilidad
Predominan en tejidos que se estiran (pulmón, arterias, piel). Ricas en Pro, Gly, Lys. Carecen de hidroxilisina.
Estructura resiliente: Moléculas de elastina (precursor tropoelastina) se unen covalentemente mediante desmosina e isodesmosina (derivados de lisina, por lisil-oxidasa), formando una red elástica 3D.
Cuando se estira, las cadenas desordenadas se ordenan; al liberar tensión, vuelven a estado relajado.
Envejecimiento: Colágeno se vuelve inflexible, elastina se degrada y no se repone → arrugas, rigidez.
Síndrome de Marfan: Mutación en Fibrilina-1 (gen FBN1), componente de microfibrillas donde se deposita la elastina. Causa sobreexpresión de TGF-β, afectando piel, pulmón, corazón, esqueleto. Peligro de ruptura aórtica. (Síntomas: extremidades largas, altura, signo de Walker-Murdoch, Steinberg).
Proteoglucanos y GAGs: Fase Gel Hidratada
Proteoglucanos: Proteína central con múltiples GAGs sulfatados. $95\%\ exto{ peso carbohidrato}.
GAGs (Glicosaminoglucanos): Polisacáridos lineales no ramificados, con cargas negativas (sulfato, carboxilo). Atraen mucha agua y cationes, creando un gel hidratado que resiste compresión y facilita difusión de moléculas.
Hialuronato (ácido hialurónico): GAG más grande y único no sulfatado ni unido covalentemente a proteína central. Sintetizado en membrana plasmática. Es lubricante (líquido sinovial) y facilita migración celular.
Condroitín sulfato, Keratán sulfato: GAGs comunes.
En cartílago, Agrecán forma agregados gigantes con hialuronato, clave para su resiliencia como "amortiguador biológico".
Funciones: Co-receptores en señalización celular (ej. Sindecanes con FGF).
Glucoproteínas Adhesivas
1. Fibronectina (FN)
Dímero grande con múltiples dominios modulares que se unen a: Colágeno, Heparina, Fibrina, y Receptores celulares (Integrinas) vía motivo RGD (Arg-Gly-Asp).
Formas: Soluble (plasma FN) en sangre (coagulación, cicatrización); Insoluble en ECM (fibrillas, guía migración celular).
Esencial para migración embrionaria. Eliminación génica letal en desarrollo (fallos cardíacos).
Cáncer: Pérdida de FN fibrilar contribuye a pérdida de adhesión y aumento de invasividad (metástasis).
2. Laminina
Principal glucoproteína adhesiva de la lámina basal (ECM especializada, aprox. ). Esta subyace a epitelios, endotelios, etc.
Heterotrímero ( cadenas) en forma de cruz asimétrica ("T").
Múltiples sitios de unión para: Colágeno IV, Heparina, Nidógeno (puente), y Receptores celulares (Integrinas: ).
Lámina Basal: Red compleja de Colágeno IV, Laminina, Proteoglucanos (Perlecán), Nidógeno.
Funciones: Soporte estructural, Filtración y barrera (ej. barrera glomerular renal), guía migración celular.
Cáncer: MMPs (Metaloproteinasas de Matriz) degradan lámina basal (colágeno IV, laminina), facilitando invasión y metástasis. MMPs también en infiltración leucocitaria.
Integrinas: Receptores Mecano-químicos
Principal familia de receptores transmembrana para adhesión célula-ECM. Funciona como heterodímeros no covalentes ( y subunidades). Ejemplos: (fibronectina), (laminina).
Dominio extracelular: se une a ECM (ej. motivo RGD).
Dominio citosólico: Se une a proteínas adaptadoras que conectan al citoesqueleto, sin actividad enzimática intrínseca. Son Calcio y Magnesio dependientes.
Conexiones Citosqueléticas:
Adhesiones focales: En células migratorias (fibroblastos). Integrinas se unen a ECM, y en citosol a talina, vinculina, -actinina, conectando a filamentos de actina. Esenciales para motilidad y señalización.
Hemidesmosomas: En células epiteliales basales. Integrina específica se une a laminina. En citosol, plectina y BPAG (antígeno de pénfigo bulloso) ligan filamentos intermedios (queratina). Adhesión fuerte, estática.
Señalización bidireccional:
"Inside-out" signaling: Señales intracelulares cambian afinidad/agrupamiento de integrinas.
"Outside-in" signaling: Unión a ECM activa quinasas (ej. FAK (quinasa de adhesión focal), ILK (quinasa ligada a integrina)), afectando proliferación, supervivencia, migración.
Crecimiento dependiente de anclaje: Células normales necesitan adhesión para sobrevivir. La falta de adhesión induce anoikis (apoptosis por falta de anclaje). Células cancerosas evaden esto.
Costameros y Distrofia Muscular
Costamero: Zonas de adhesión en membrana muscular, conectan miofibrillas a ECM. Actúa como "ancla" y "amortiguador".
Complejo distrofina/distroglicano: Enlace clave.
Distrofina: Proteína citosólica, une filamentos de actina.
Distroglicanos (): Proteínas transmembrana, unen distrofina a Laminina (Laminina-211) en lámina basal muscular.
Distrofias Musculares de Duchenne (DMD) y Becker (BMD): Causadas por mutaciones en gen de la distrofina. Proteína ausente o disfuncional interrumpe conexión → debilidad muscular progresiva, daño crónico, falla respiratoria/cardíaca.
Cultivos 3D, Matrigel y Bioingeniería (Key Technique)
Problema: Cultivos 2D no replican ECM in vivo.
Solución: Matrigel (extracto de lámina basal de tumor). Permite células crecer en 3D.
Ejemplo: Células epiteliales mamarias (MCF-10A) forman quistes polarizados en Matrigel, imitando estructuras glandulares.
Importancia: Demuestra control de ECM sobre forma, diferenciación, proliferación y comportamiento maligno. Crucial en ingeniería tisular (órganos de laboratorio, regeneración de tejidos).
Pared Celular Vegetal: Analogía y Diferencias
Estructura extracelular rígida en plantas; provee esqueleto externo, resiste presión de turgencia, barrera protectora.
Composición: Celulosa (microfibrillas, resistencia a tensión, ), Hemicelulosas (), Pectinas (matriz gelatinosa, resistencia a compresión, ), Glucoproteínas (Extensinas), Lignina (en madera).
Arquitectura: Como hormigón armado (Celulosa = varillas de acero; Hemicelulosas/Extensinas = red; Pectina = matriz gelatinosa).
Comparación con ECM animal: Ambos soporte, pero: ECM animal (colágeno, elastina, proteoglucanos, fibronectina/laminina); Pared vegetal (celulosa, hemicelulosas/extensinas, pectinas).
Implicaciones Filosóficas y Éticas
Destino celular: Influenciado por contexto ambiental (ECM) no solo por genes.
MMPs: Degradación excesiva en cáncer (dilema terapéutico: inhibir MMPs sin afectar funciones fisiológicas).
Ingeniería tisular: Debates bioéticos sobre definición de vida, modificación organismos, equidad en acceso.