Lecture 17 - MHC
TCR and Antigen Recognition
TCRs recognize peptide fragments
TCR has a single peptide binding site
the TCR MHC must also contact through two connection points
MHC molecule structure and classes
MHC: major histocompatibility complex (HLA)
membrane bound glycoproteins split into class I and class II
Class I MHC
alpha chain with three domains and a non-covalently associated B2 microglobulin
microglobulin in a small Ig-like domain
only the alpha chain forms the binding cleft
Class II MHC
alpha and beta chain with two domains each, non-identical
both form the binding cleft
peptide loading versatility and constraints
MHC have the capacity to bind a wide variety of peptide sequences
only one peptide can bind to each MHC protein at a time
peptides fit into binding cleft of MHC molecules
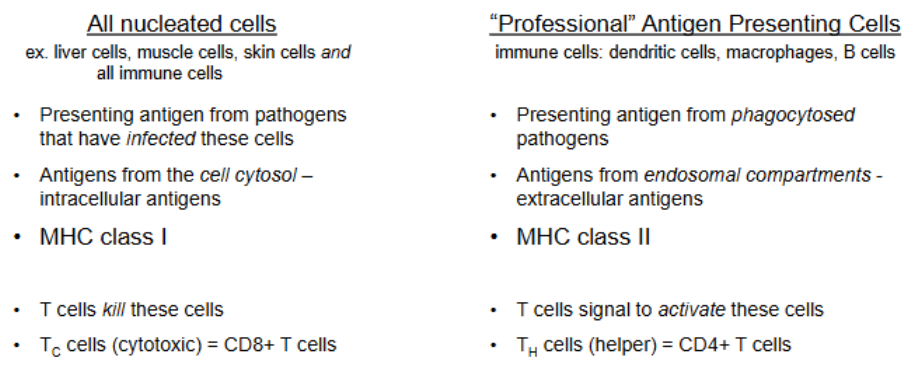
Recap on pathogen recognition
pathogens can be found in the cytosol or in endosomal/phagocytic compartments
cytosolic — MHC I (all nucleated cells: “I am sick”)
endosome — MHC II (professional APCs: “I need help”)
if the pathogen escapes into the cytosol, the cell utilizes MHC I
MHC class I and II expression
class II expressed by professional APCs
B cells, macrophages, and dendritic cells
class I expressed by most nucleated cells
T cells, B cells, macrophages, dendritic cells, neutrophils, hepatocytes, kidney epithelium, and neurons
MHC :: T cell co-receptor interactions
CD8 — Cytotoxic T cell — MHC I class — cytosol
release of perforin and granzymes to induce apoptosis of the infected cell
CD4 — Helper T cells — MHC II class — phagosome
secretion of cytokines to instruct macrophages to enhance killing and B cells to produce antibodies
Endomembrane system foundations for antigen processing
ribosomes exist freely in the cytoplasm or on the rough ER
both MHC I and II are synthesized within the ER by ribosomes
inside the ER, they fold and are membrane-bound
they are packaged into vesicles that are delivered to the G.
depending on the tag, vesicles are shipped to different parts of the cell or to the outside
endocytosis brings in outside material
the endosome progresses through early to late endosome phase
phagocytosis generates phagosome which fuse with lysosomes that contain enzymes activated at low pH (get low pH by proton pumps)
cellular compartmentalization
three broad compartments: nucleus & cytoplasm, endomembrane system, and mitochondria
MHC Class I antigen processing pathway
stage one — degradation of proteins by the proteosome
cytosolic proteins are tagged by ubiquitination (labels protein to become targeted for degradation)
the proteins are degraded by proteosome into peptide fragments
constitutive proteasome (baseline)
immunoproteasome (induced by IFN-gamma expression to generate peptides more suitable for MHC I)
increases the rate of degradation
stage two — import of peptides into the ER
TAP is a transporter associated with antigen processing
TAP brings peptides from the cytosol into the ER lumen
stage three — binding of peptides to empty MHC class I proteins in the ER
ERAP trims overlong peptides to appropriate size for the MHC-I groove
membrane bound calnexin and other proteins hold the MHC I into an inactive configuration
TAP recruitment and associated assembly factors displace calnexin and facilitate peptide loading
stage four — transport of loaded MHC to the cell surface
after peptide loading, MHC-I is packaged in vesicles, passes through Golgi, and traffics to the plasma membrane for presentation to CD8+ T cells
How cells distinguish viral vs. self proteins (rationale for degradation bias)
Increased error/misfolding under viral-driven ramp-up of protein synthesis leads to misfolded proteins that are ubiquitinated and degraded regardless of origin
Post-translational modification deficits: viral proteins often lack normal host signals for modifications (e.g., addition of lipids/sugars), flagging them as abnormal
Trafficking address/tag deficits: viral proteins often lack proper localization signals, making them “missing address” proteins; cells default to degrade and recycle such proteins
General principle: although amino acid composition overlaps, quality control and signaling deficiencies in viral proteins bias them toward ubiquitination and degradation
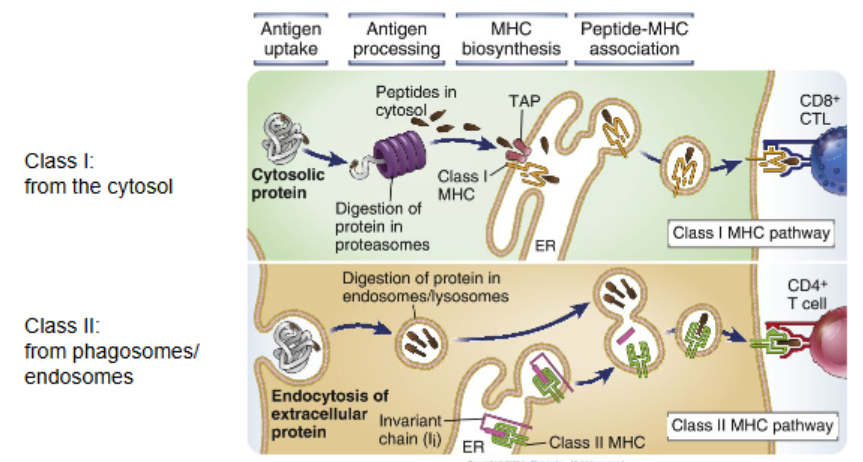
MHC Class II antigen processing pathway
stage one — degradation of pathogen in phagolysome
encounter of antigen and consumption via endocytosis/phagocytosis
these vesicles fuse with lysosomes to yield fragmented peptides of the antigen
stage two — fusion of endosomal compartment containing MHC class II in the membrane
MHC class II molecule is made in the rough ER; it has a small fragment invariant chain bound to prevent unnecessary binding
MHC-II with invariant chain is trafficked via Golgi into vesicles targeted to the endosomal/lysosomal system
stage three — binding of peptides to empty MHC class II
fusion of MHC-II–containing vesicles with peptide-rich endolysosomal compartments results in removal of the invariant chain and replacement by exogenous peptides processed in the acidic lumen
stage four — transport of loaded MHC to the cell surface
loaded MHC-II is then presented at the plasma membrane to CD4+ helper T cells
Antigen-crossing presenting
self-peptides are commonly encountered during a T cell’s maturation phase
undergoes positive and negative selection to ensure there is no response to self cells
to ensure that infection at any location throughout the body can be detected
phagocytosis and then dendritic cells present antigens on MHC class I to activate CD8+ T cells which kill infected cells
MHC Diversity
multiple class I and II genes (isotypes)
class isotypes of MHC I molecules
HLA-A, HLA-B, HLA-C, HLA-E, HLA-F, and HLA-G
class isotypes of MHC II molecules
HLA-DP, HLA-DR, HLA-DO, HLA-DQ, and HLA-DM
mix and match between alpha/beta chains for MHC II molecules
co-dominant expression of all MHC molecules
many different isotypes with their own promiscuity allow cells to display a large number of peptides
MHC diversity allows for presentation of a wide range of peptides
binding pocket to accommodate proteins & two amino acids of the peptide interact with the binding cleft
those two amino acids must be kept the same but the other resides can be variable
genetic polymorphism within these genes (allotypes
polymorphisms primarily occur in the peptide-binding groove
diversity protects human population
MHC genes are linked: crossover limited
offspring inherit a ‘set’ of alleles either from the mother or father
crossover limited events still generate HLA polymorphism