Fatty Acid Breakdown
Fat Metabolism and Adipose Tissue Characteristics
Fat serves as the primary energy storage mechanism in mammals.
half of our ATP comes from fatty acid breakdown at rest
After our Glucose energy store is finished we rely of fatty acid breakdown.
Quantitative Storage: In a non-obese person weighing , there is approximately of stored triacylglycerol (triglyceride).
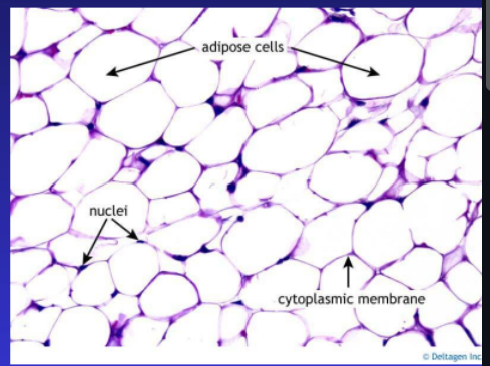
Adipose Tissue: Is a type of connective tissue in the body that primarily stores fat
Appears as globules within adipocytes (adipose cells).
These cells contain nuclei and a cytoplasmic membrane.
in adipose tissue fatty acids are stored as Triacylglycerol (triglyceride)
these r both the same thing
Nature of Fat:
Highly Concentrated
Stores more energy in a smaller volume than other macronutrients.
Anhydrous
It is stored without water, unlike glycogen which is hydrated.
Highly Reduced
Fatty acids contain a high proportion of carbon-hydrogen bonds.
so when oxidised you get lots of energy
contains less oxygen
Energy Density:
Fat yields significantly higher energy per gram compared to carbohydrates (glucose/glycogen).
Structure of Triacylglycerols and Fatty Acids
What is the main energy store in the body
glycogen
stored in liver and skeletal muscles
Why isn’t glycogen the main energy sourced used
glycogen has a lot of water in it.
where as Triacylglycerol doesn’t
so you can store more of it
Triacylglycerol (TAG) Composition
consists of a glycerol backbone that is esterified with three fatty acid chains
Classification of Fatty Acids:
Saturated: No double bonds (e.g., Stearic acid).
Monounsaturated: One double bond (e.g., Oleic acid).
Polyunsaturated: Multiple double bonds (e.g., Linoleic acid).
Chemical Formulas and Specific Examples:
Palmitate:
Linoleate:
Mechanism of Triacylglycerol Breakdown
General Products
free fatty acids
glycerol.
Key Enzyme:
Hormone-sensitive lipase (also known as triacylglycerol lipase) mediates this process.
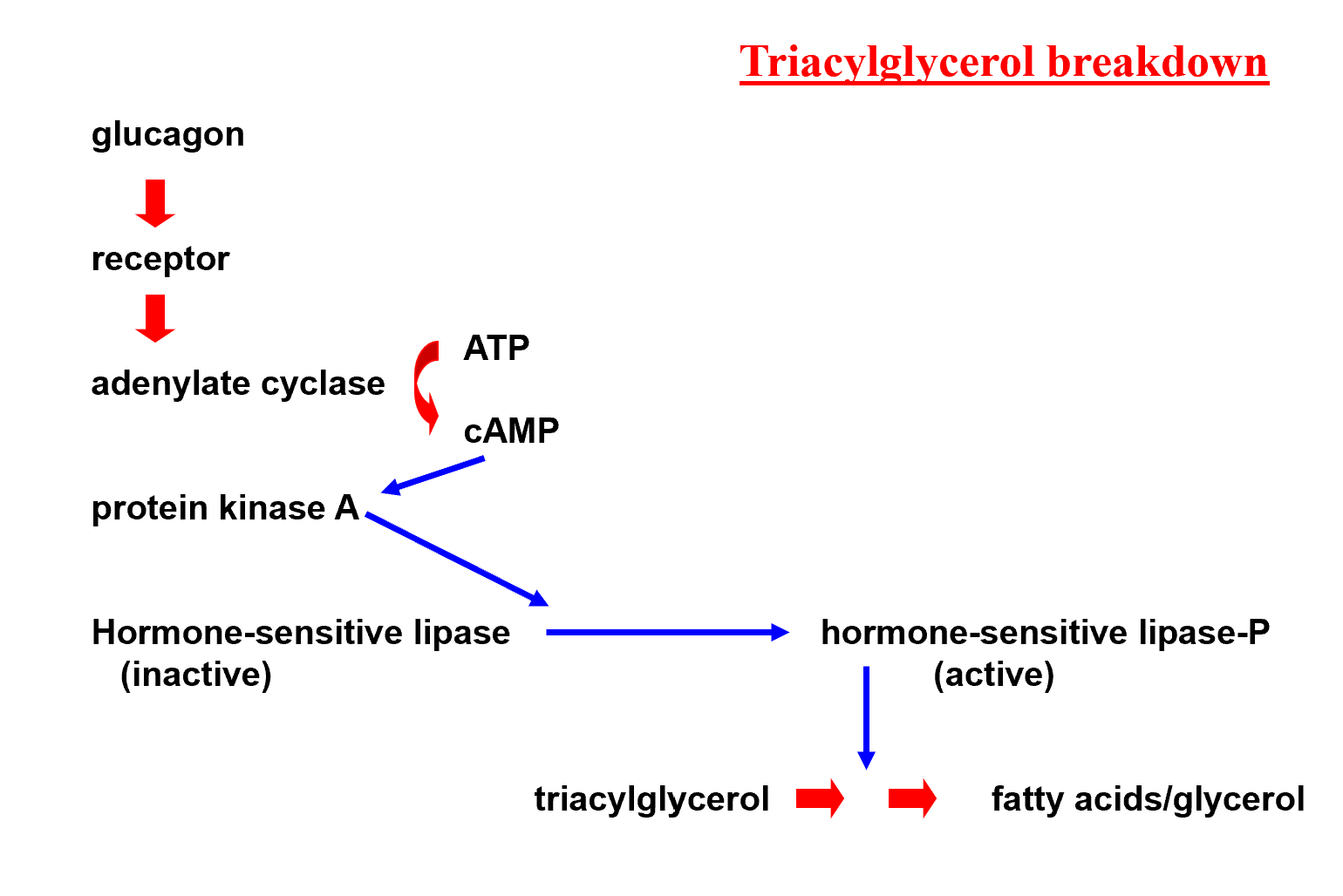
Hormonal Stimulation:
The process is stimulated by catabolic hormones such as glucagon and adrenaline (epinephrine).
Signaling Pathway and Control:
1. Hormone Binding:
Glucagon binds to its specific receptor on the cell membrane.
2. Activation of Adenylate Cyclase
This activation catalyzes the conversion of into .
3. Protein Kinase A (PKA) Activation
Increased levels of activate Protein Kinase A.
4. Enzyme Phosphorylation
PKA phosphorylates the inactive Hormone-sensitive lipase, converting it into active Hormone-sensitive lipase-P.
5. Lipolysis
Active Hormone-sensitive lipase breaks down triacylglycerols into fatty acids and glycerol.
Metabolic Fate of Products:
Fatty Acids: Utilized as fuel through -oxidation.
Glycerol: Utilized in glycolysis or gluconeogenesis.
Fatty Acid Transport and Circulation
Water Insolubility:
Fatty acids are insoluble in water and cannot travel freely in the aqueous environment of the blood plasma.
so instead they bind to albumin
Albumin Binding:
Upon release from adipose tissue, fatty acids bind to albumin in the plasma for circulation.
albumin is the most abundant protein in plasma
Distribution: They are transported to high-energy-demand tissues such as skeletal muscle or the heart for -oxidation.
this is when 2 carbon units removed in each cycle
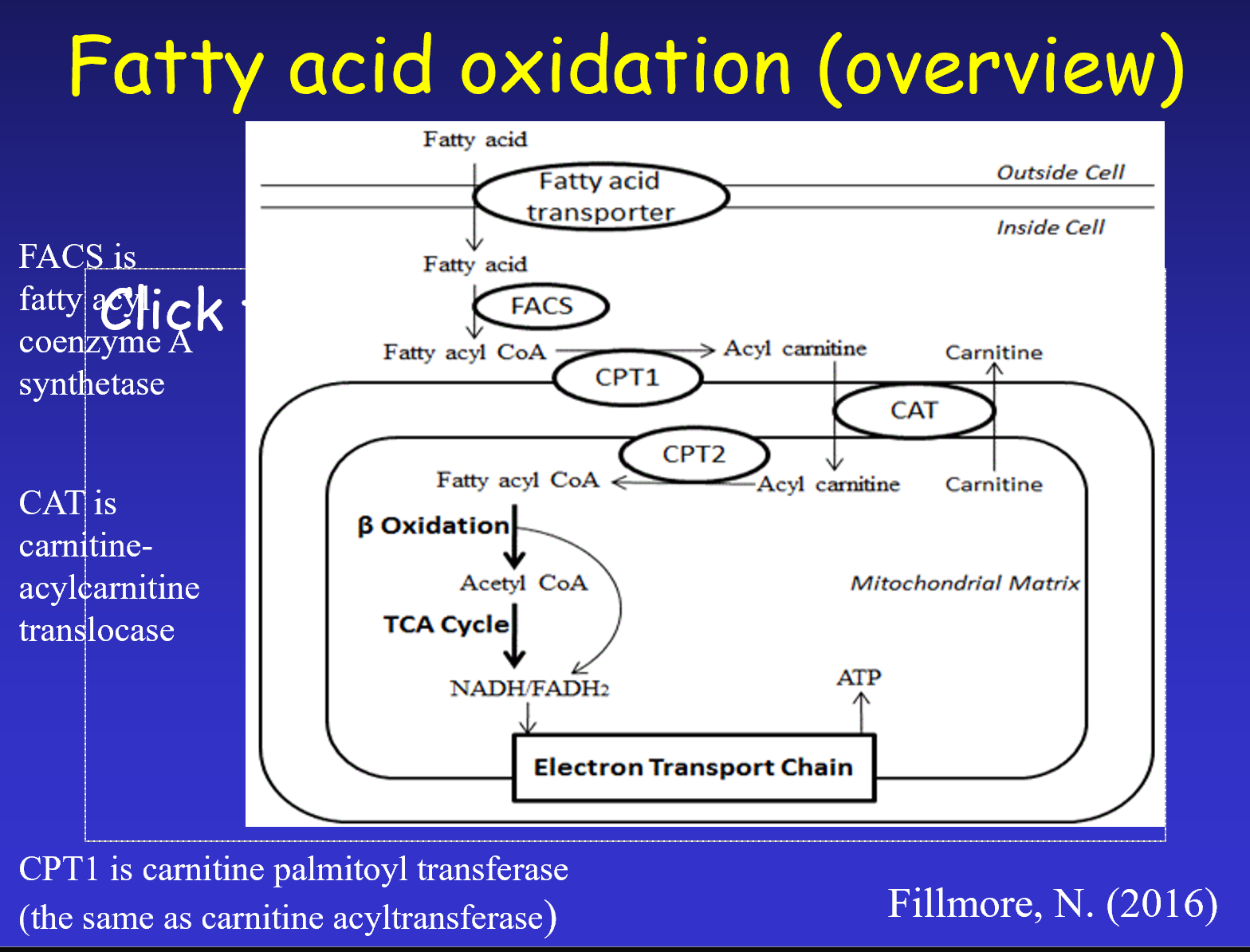
Entry into the Mitochondrion
We need to get fatty acids from the cytosol into the mitochondria
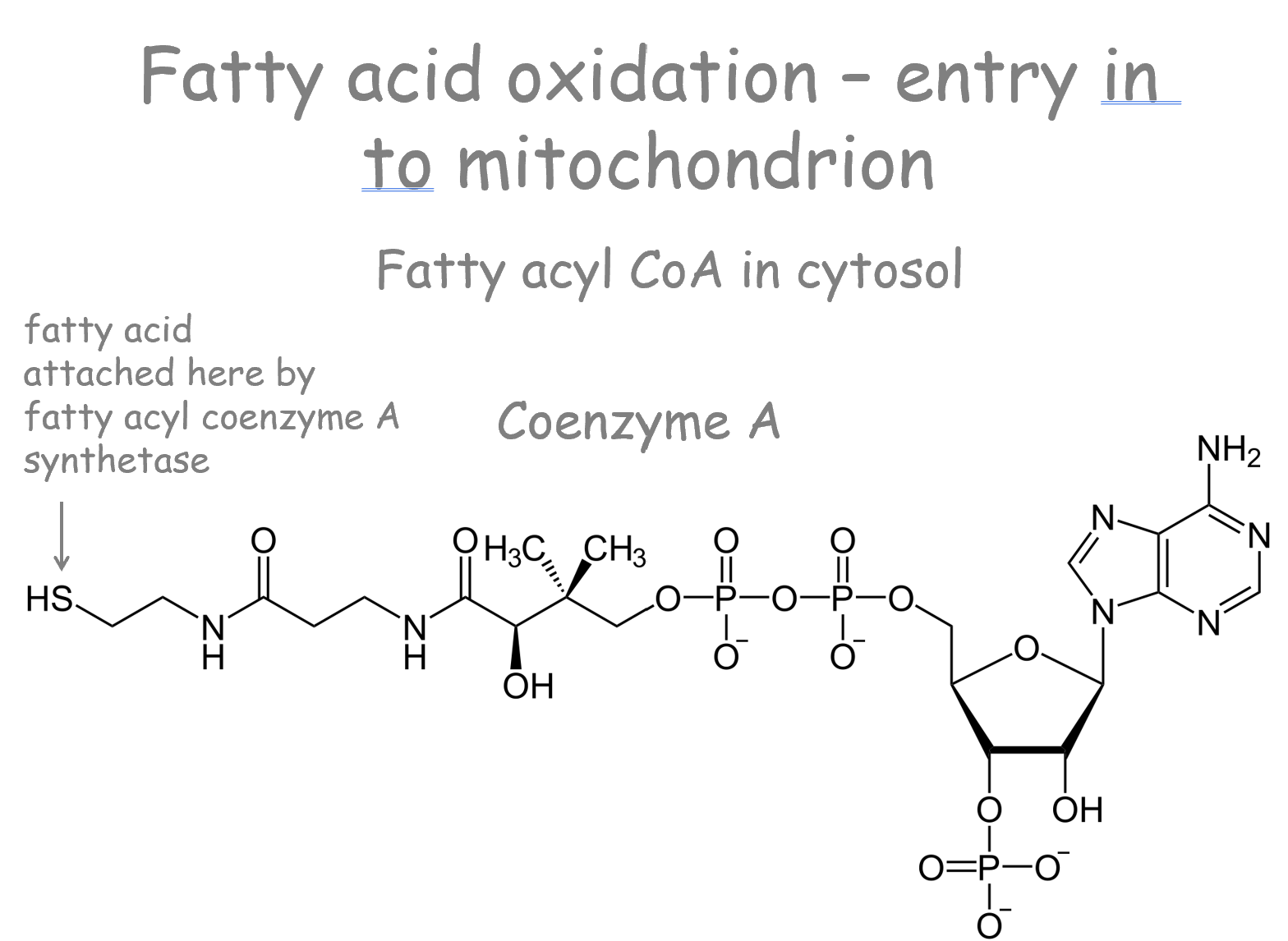
Activation in Cytosol:
Fatty acids are first attached to Coenzyme A
by the enzyme fatty acyl coenzyme A synthetase (FACS)
forming Fatty acyl CoA.
Membrane Permeability: The outer mitochondrial membrane has pores and is permeable. However, the inner mitochondrial membrane (IMM) is impermeable to long-chain fatty acyl CoAs.
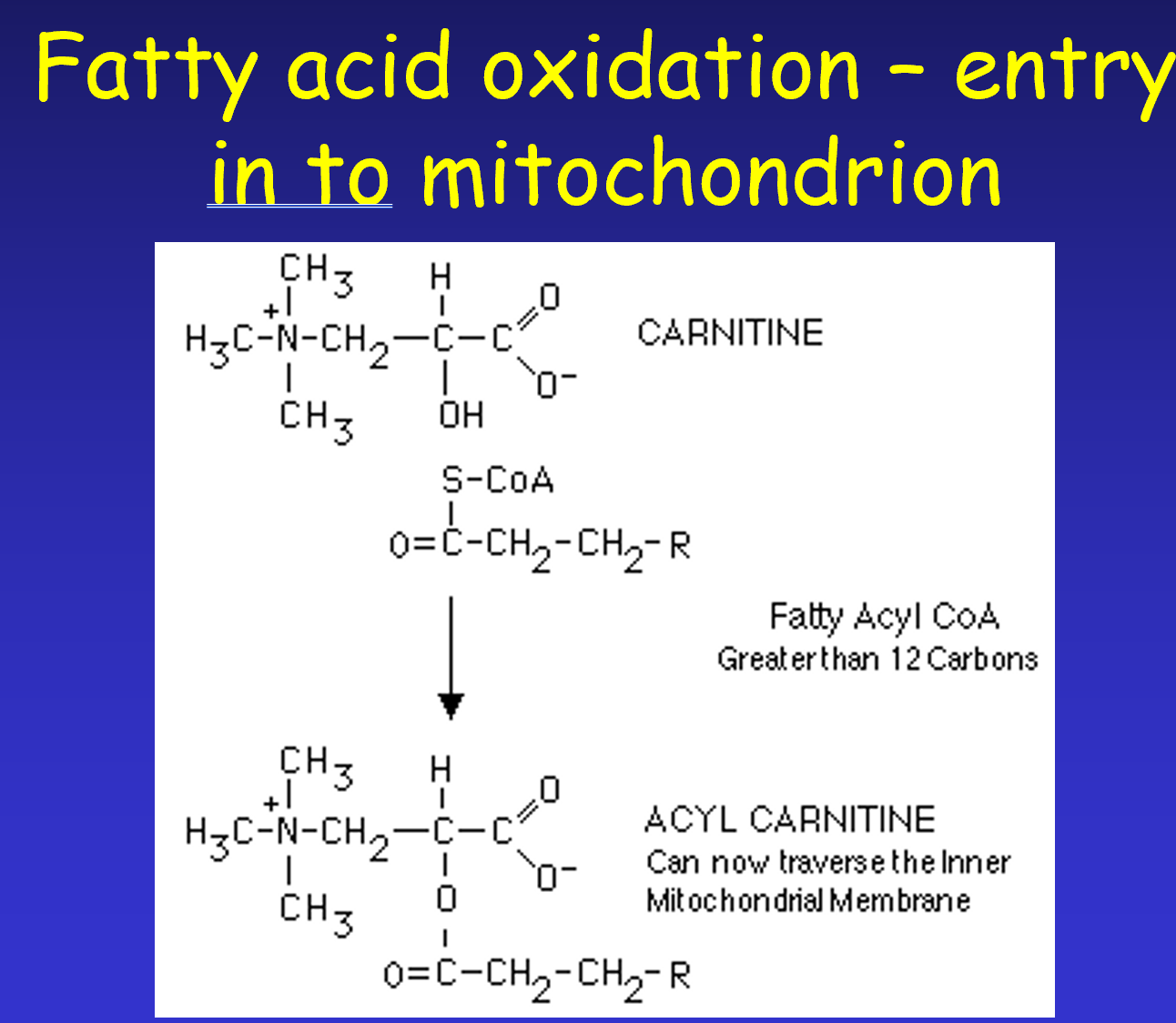
The Carnitine Shuttle:
Threshold: Fatty acids with more than carbons require a transport mechanism.
Conversion: Fatty acyl CoA in the cytosol is converted to acyl carnitine by the enzyme carnitine acyltransferase (also called carnitine palmitoyl transferase 1 or CPT1).
Translocation: Acyl carnitine traverses the IMM via a translocase called carnitine-acylcarnitine translocase (CAT).
Reconversion: Once inside the mitochondrial matrix, acyl carnitine is reconverted back into fatty acyl CoA.
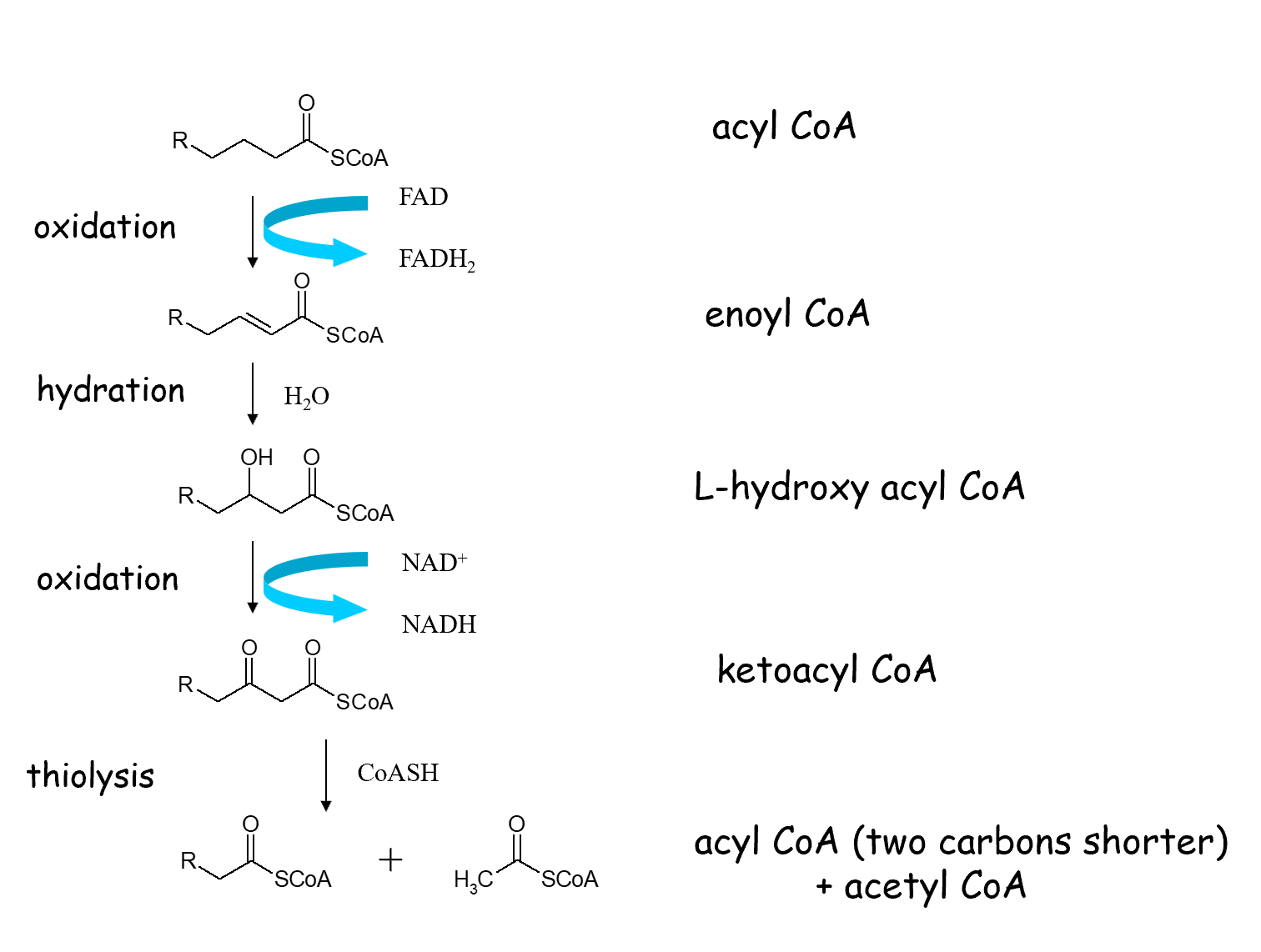
Mitochondrial -Oxidation Cycle
Process Overview: Fatty acid oxidation occurs in the mitochondria and removes two-carbon units in each sequential cycle.
The Four Steps of -Oxidation:
1. Oxidation: Acyl CoA is oxidized to Enoyl CoA, reducing to .
2. Hydration: Enoyl CoA is hydrated with to form L-hydroxy acyl CoA.
3. Oxidation: L-hydroxy acyl CoA is oxidized to ketoacyl CoA, reducing to .
4. Thiolysis: Ketoacyl CoA reacts with Coenzyme A () to form Acetyl CoA and a Fatty acyl CoA that is two carbons shorter than the original.
Energy Yield from Palmitoyl CoA:
Complete oxidation of one palmitoyl CoA molecule ultimately yields ATP.
This process captures approximately of the available energy from the fatty acid.
the rest of the energy goes to heat
Specific Cases and Regulation of Fatty Acid Oxidation
Odd-Numbered Fatty Acids:
Found in small amounts in some vegetables.
Their oxidation yields Acetyl CoA and one molecule of Propionyl CoA.
Propionyl CoA is subsequently converted to Succinyl CoA to enter the citric acid cycle.
Unsaturated Fatty Acids: b Oxidation requires auxiliary enzymes to move double bonds from one position to another or to remove double bonds by making them suitable for the standard cycle.
Control Mechanisms:
Supply: Regulated by the rate of triacylglycerol breakdown in adipose tissue.
Inhibition by Malonyl CoA: Malonyl CoA, an intermediate in fatty acid synthesis, inhibits carnitine acyltransferase (acyl carnitine production).
This prevents a futile cycle by stopping fatty acid breakdown when fuel molecules (and thus fatty acid synthesis) are abundant.
Ketone Bodies
Ketone bodies: Small water‑soluble molecules made in the liver from fatty acids, and they act as an alternative fuel source when glucose is low.
Synthesis Conditions:
Ketone bodies are synthesized in the liver when Acetyl CoA levels are high but oxaloacetate levels are insufficient to maintain the citric acid cycle.
Example Scenarios: Diabetes and starvation.
Mechanism: Fatty acids from adipose tissue are oxidized to form Acetyl CoA in the liver. However, if oxaloacetate is being diverted for gluconeogenesis, its concentration falls. Without enough oxaloacetate to combine with Acetyl CoA to form citrate, Acetyl CoA is diverted to ketone body synthesis.
When fatty acids are broken down (β‑oxidation):
Lots of acetyl‑CoA is produced
But the Krebs cycle slows (because oxaloacetate is used for glucose production)
👉 So acetyl‑CoA builds up
The liver solves this by converting excess acetyl‑CoA into ketone bodies
Types of Ketone Bodies:
Acetoacetate - gluconeogenesis
Acetone
-hydroxybutyrate (or -hydroxybutyrate)
Physiological Role:
Solubility: Ketone bodies are water-soluble.
Fuel Source: Heart, kidney cortex, and brain.
Brain Metabolism: Normally uses glucose, but uses ketone bodies during starvation. The brain cannot use fatty acids directly because they cannot cross the blood-brain barrier.
They are released into the blood and used by other tissues:
✅ Brain (during prolonged fasting)
✅ Muscle
✅ Heart
⚠ The liver itself cannot use ketone bodies
Ketoacidosis: Because ketone bodies (like acetoacetate and $\beta$-hydroxybutyrate) are acidic, excessive production can lead to ketoacidosis.
Comparative Metabolism: Animals vs. Plants
Limitations in Animals:
Animals can enter Acetyl CoA into the citric acid cycle to produce .
The Pyruvate Dehydrogenase reaction (Pyruvate to Acetyl CoA) is irreversible.
Consequently, animals cannot convert Acetyl CoA into glucose, meaning they cannot convert fatty acids into glucose.
The Glyoxylate Cycle in Plants and Bacteria:
Plants and bacteria possess the glyoxylate cycle, allowing them to convert Acetyl CoA into oxaloacetate.
This bypasses the steps of the citric acid cycle that release , facilitating glucose synthesis from fatty acids.
Significance: This is prominent in oil-rich seeds (e.g., sunflowers), allowing seedlings to grow using stored fat until they can perform photosynthesis.
Organelle: This cycle occurs in specialized organelles called glyoxysomes.