13. Antibiotic Production III-> Biosynthesis
A reminder: how are natural product antibiotics synthesised?
Dedicated biosynthetic pathways exist in bacteria and fungi for the production of antibiotics (as well as compounds with other activities)
Huge variety of pathway types - often very complex
Genes for these pathways are almost always clustered in bacteria and fungi
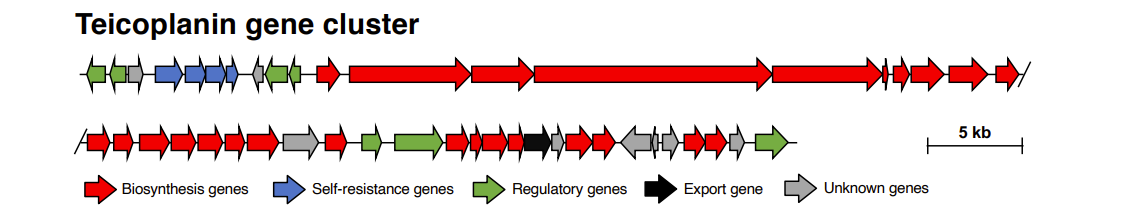 Gene cluster encodes proteins for the teicoplanin regulation, biosynthesis, self-resistance and export
Gene cluster encodes proteins for the teicoplanin regulation, biosynthesis, self-resistance and export
Non-ribosomal peptides
Common features
Peptide backbone
Non-proteinogenic amino acids
Oxidative cross links
D-amino acids
Cyclisation
Acylation and glycosylation
Elucidating Non-Ribosomal Peptide Biosynthesis: Penicillin
Process of understanding the mechanism by which non-ribosomal peptides, such as penicillin, are synthesized in organisms.
Non-ribosomal peptide biosynthesis is a complex pathway that involves the assembly of amino acids into peptides without the involvement of ribosomes. Penicillin, a well-known antibiotic, is produced through non-ribosomal peptide biosynthesis in certain fungi and bacteria.
The elucidation of this process involves studying the enzymes, genes, and regulatory mechanisms involved in the synthesis of non-ribosomal peptides like penicillin.
By understanding this biosynthetic pathway, scientists can potentially manipulate it to produce novel antibiotics or other bioactive compounds.
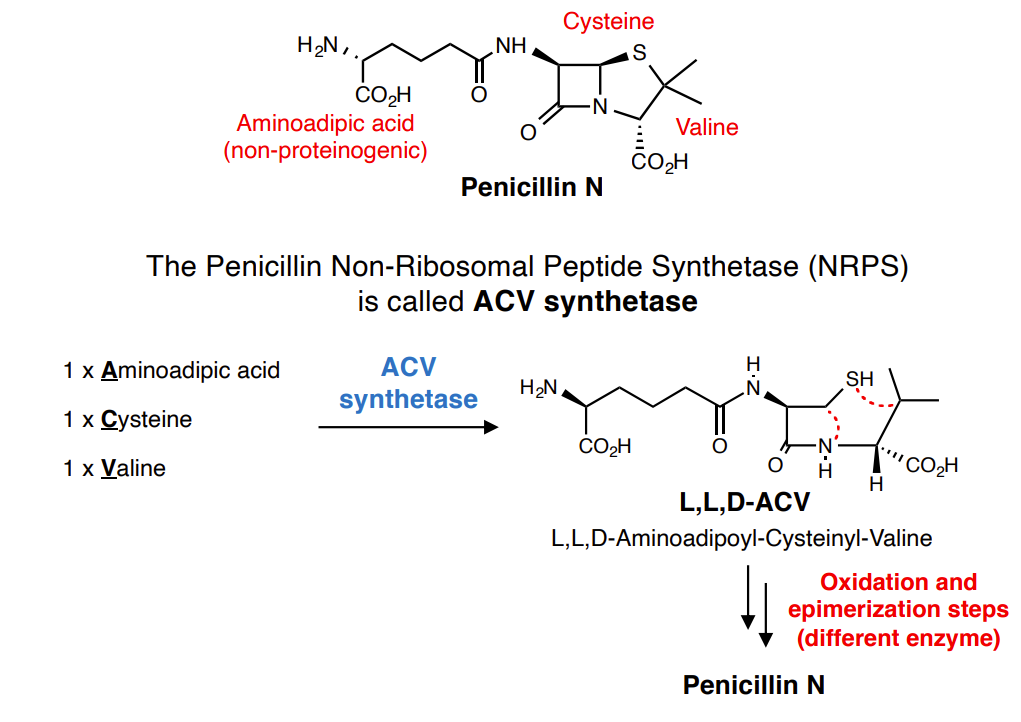 The Penicillin Non-Ribosomal Peptide Synthetase (NRPS)
The Penicillin Non-Ribosomal Peptide Synthetase (NRPS)
ACV synthetase is a massive multi-domain protein
Each module incorporates one amino acid
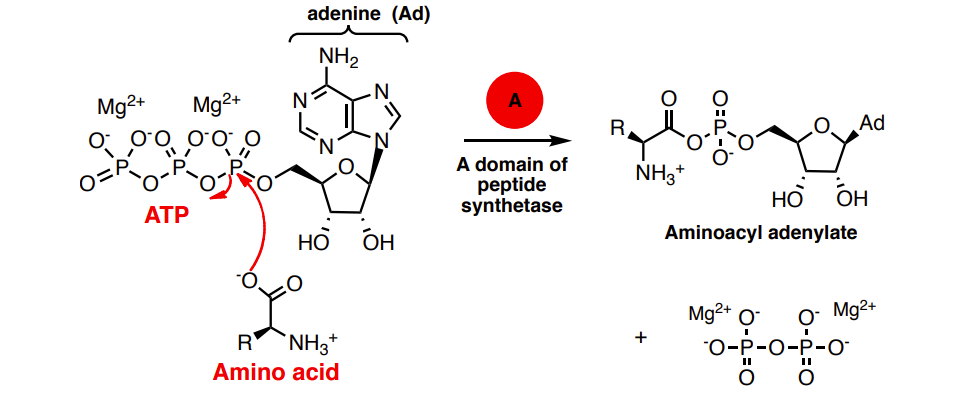
 Adenylation (A) domain Activates amino acids as aminoacyl adenylates using ATP
Adenylation (A) domain Activates amino acids as aminoacyl adenylates using ATP
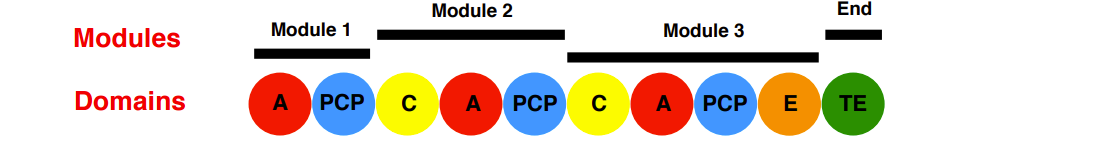
 The Penicillin Non-Ribosomal Peptide Synthetase (NRPS)
The Penicillin Non-Ribosomal Peptide Synthetase (NRPS)
Substrate tethering: The Thiolation (T) Domain or Peptidyl Carrier Protein (PCP)
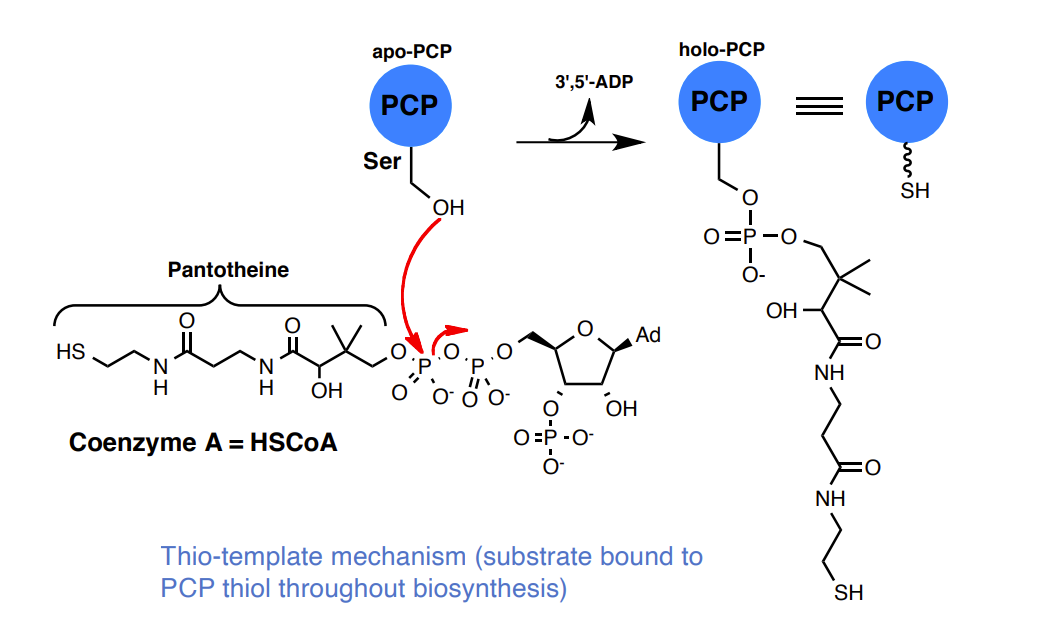 Interaction between the A and PCP domains: aminoacyl thioester formation
Interaction between the A and PCP domains: aminoacyl thioester formation
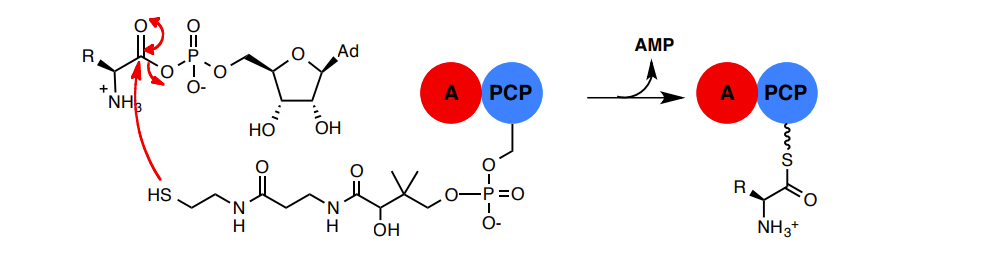 Amide bond formation: The Condensation (C) domain
Amide bond formation: The Condensation (C) domain
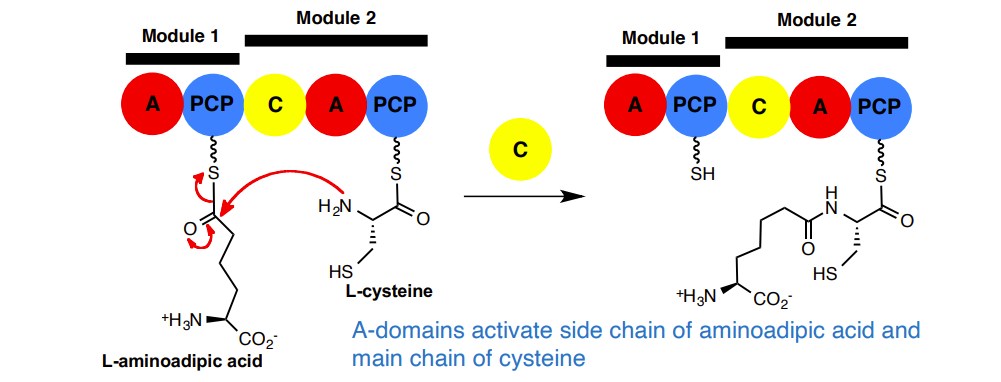 A-domains activate side chain of aminoadipic acid and main chain of cysteine
A-domains activate side chain of aminoadipic acid and main chain of cysteine
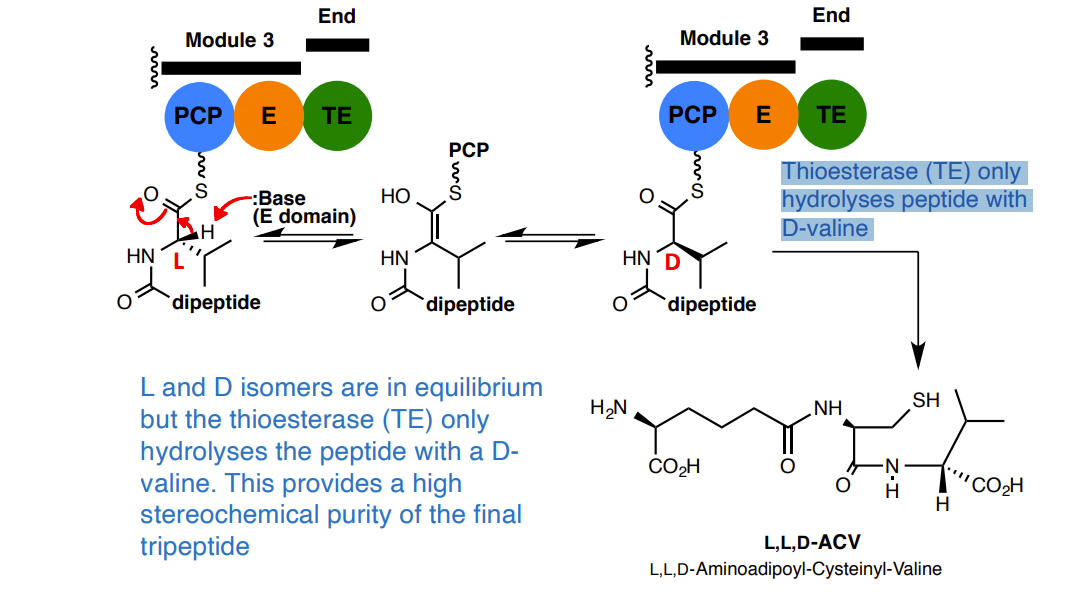
Incorporation of the D-valine residue
1. Activation and condensation of L-valine
D-valine is not incorporated
Instead, adenylation (A) and condensation (C) domains select for L-valine
How does D- stereochemistry get introduced at this position?
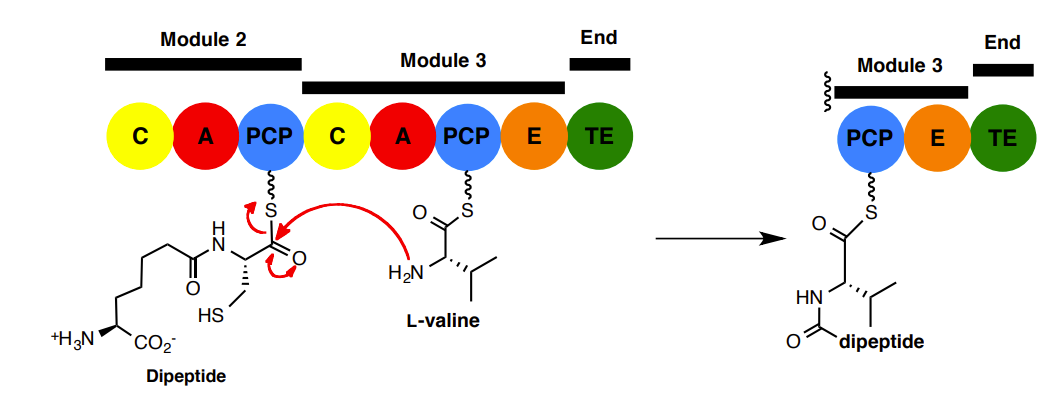 2. The Epimerisation (E) domain
2. The Epimerisation (E) domain
L and D isomers are in equilibrium but the thioesterase (TE) only hydrolyses the peptide with a D-valine. This provides a high stereochemical purity of the final tripeptide
Thioesterase (TE) only hydrolyses peptide with D-valine
 ACV Synthetase Summary
ACV Synthetase Summary
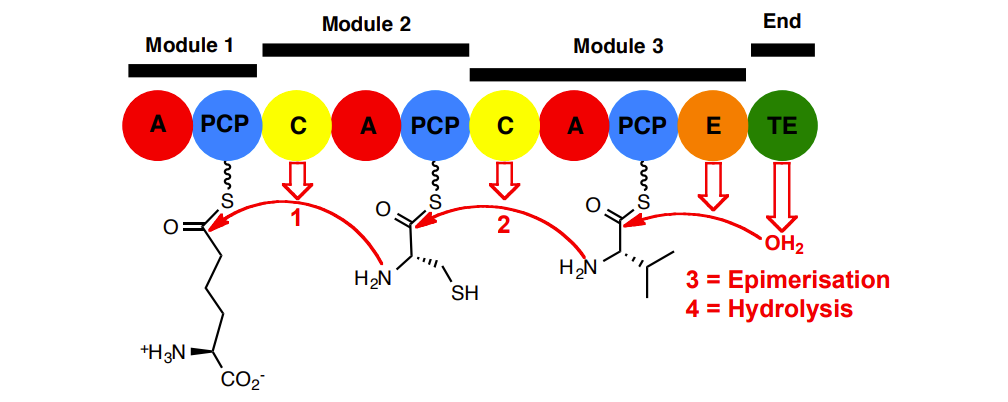
A domains have specificity for loading correct amino acid onto the adjacent PCP (sometimes called a T domain).
C domains catalyse the peptide bond forming step. They have a substrate specificity for the correct enantiomers.
E domains catalyse epimerisation of the peptidyl-thioester
TE domain catalyses the off-loading hydrolysis (cyclisation in some other NRPSs). It too is stereoselective.
Multiple enzymatic proof-reading steps ensure product (stereo)chemical purity
Further NRPS domains to introduce diversity
E → Epimerisation domain: Converts residue from L- to D-amino acid. Structurally related to C domains.
C/E → Dual Condensation/Epimerization domain: Enables D-amino acid incorporation without a separate E domain.
Cy → Cyclisation domain: Catalyses condensation and then heterocyclisation of cysteine, serine, or threonine residues
Ox → Oxidase domain: Oxidation of cyclised Cys/Ser/Thr residues
MT → Methyltransferase domain: Methylation of backbone amide nitrogen
R → Reductase domain: Present instead of thioesterase (TE) domain. Catalyses the two- or four-electron reductive off-loading of a peptidyl thioester to a C-terminal aldehyde or alcohol
Presence of one or more of these domains (e.g., E domain in penicillin biosynthesis) provides additional structural diversity to the resulting natural product
Discovery of nisin
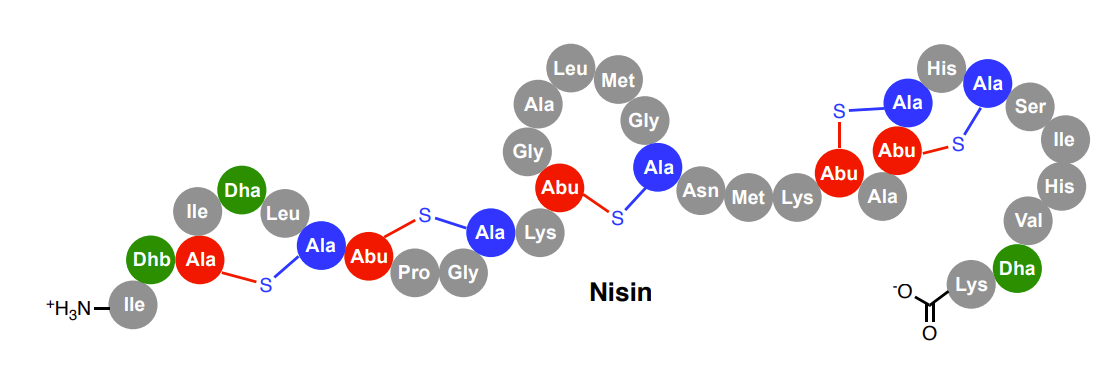
Produced by Lactococcus lactis
Broad bactericidal activity towards Gram-positive bacteria
Used for over 50 years as a food preservative (E234)
Antibacterial activity against multiple clinically important pathogens such as MRSA, VRE and Streptococcus pneumoniae
Huge molecule for a natural product (3.3 kDa)
Inhibits cell wall biosynthesis and generates pores in bacterial membranes
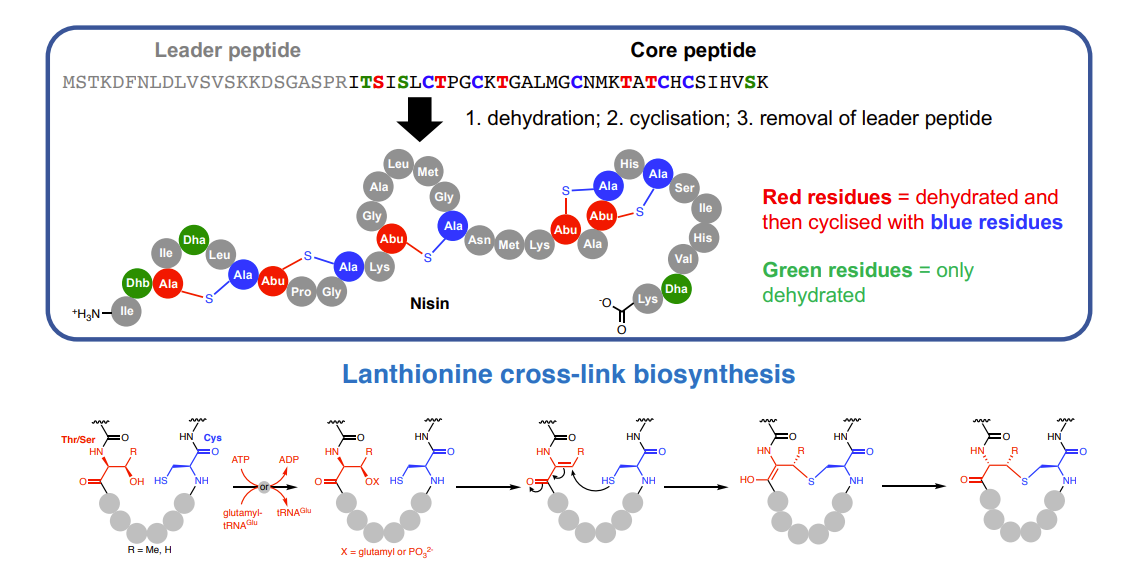
Biosynthesis of nisin
Belongs to a RiPP family called lanthipeptides
Antibacterial lanthipeptides are called lantibiotics
Lanthionine cross-links within the peptide provide stability towards proteases
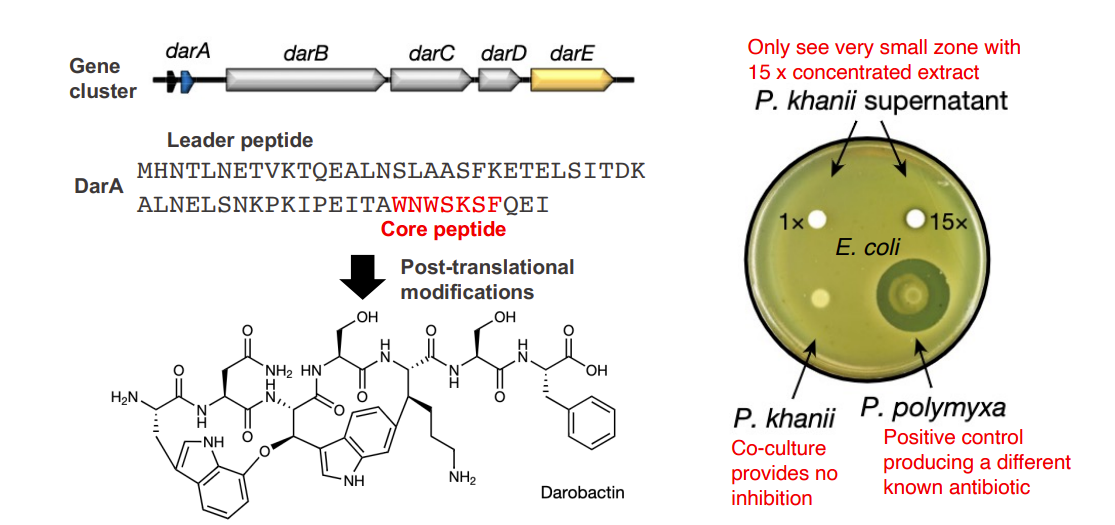
Discovery of a new RiPP antibiotic candidate
Screened a set of Photorhabdus and Xenorhabdus strains against E. coli
A (nearly) “silent” pathway in Photorhabdus khaini
Only see activity with a highly concentrated extract due to poor expression of pathway
Potent activity towards Gram-negative pathogens but not Gram-positive bacterica
Engineering RiPP antibiotics
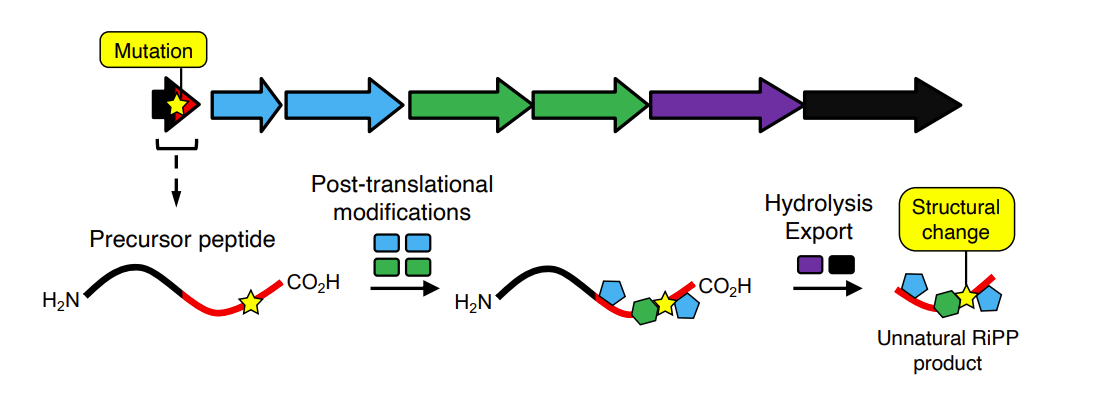
Ribosomal origin enables rapid modification of RiPP product via mutations to precursor gene
Can generate large libraries of derivatives or make targeted mutations
Requires pathway to tolerate mutations during processing of peptide into final molecule - is quite common to have good tolerance to mutations but does vary
Target is BamA, a chaperone that catalyses folding and insertion of new β-barrel proteins into the bacterial outer membrane
Used a heterologous expression system to improve yield and engineer darobactin production
Summary across the antibiotics lectures
The discovery and development of antibiotics represented a medical revolution of the 20th century
Natural products have proven to be excellent antibiotics
Widespread multidrug resistance combined with a lack of recent discoveries means that there is a pressing need to identify new antibacterial therapies
A variety of new scientific technologies has enabled the recent discovery of many promising antimicrobials
The financial model for antibiotic drug discovery may need changing before many truly new antibiotics make it to market due to the risk associated with developing new antibiotics