23b-Biochemistry-Lecture23b | Week 13 - Lecture 1
Lipid Metabolism and Fatty Acid Biosynthesis
Key Concepts in Fatty Acid Metabolism
Pathway Distinction:
Catabolism (oxidation) of fatty acids (breakdown):
Occurs in mitochondria
Forms acetyl-CoA and produces NADH
Anabolism (synthesis):
Takes place in cytosol of animals (and chloroplasts of plants)
Requires both acetyl-CoA and malonyl-CoA as starting materials
Utilizes NADPH as reducing power
Fatty Acid Biosynthesis Steps
Fatty acids are built in several passes, adding one acetate unit at a time by the fatty acid synthase complex (in the cytoplasm, where NADPH/NADP⁺ is high).
Acetate is converted to malonyl-CoA (a 3-carbon intermediate) using acetyl-CoA.
Malonyl-CoA formation is catalyzed by Acyl CoA Carboxylase (ACC).
Each pass reduces a carbonyl carbon to a methylene carbon.
Synthesis is similar to β-oxidation in reverse, but uses:
Different enzymes
Different cofactors
Occurs in a different location
The end product is palmitoyl-CoA, which inhibits ACC (negative feedback).
Comparison: Fatty Acid Synthesis vs β-Oxidation
β-Oxidation: Repeated cycles of oxidation, hydration, oxidation, cleavage.
Synthesis: Repeated cycles of condensation, reduction, dehydration, reduction.
Opposite reactions but with different enzymes and energy flow.
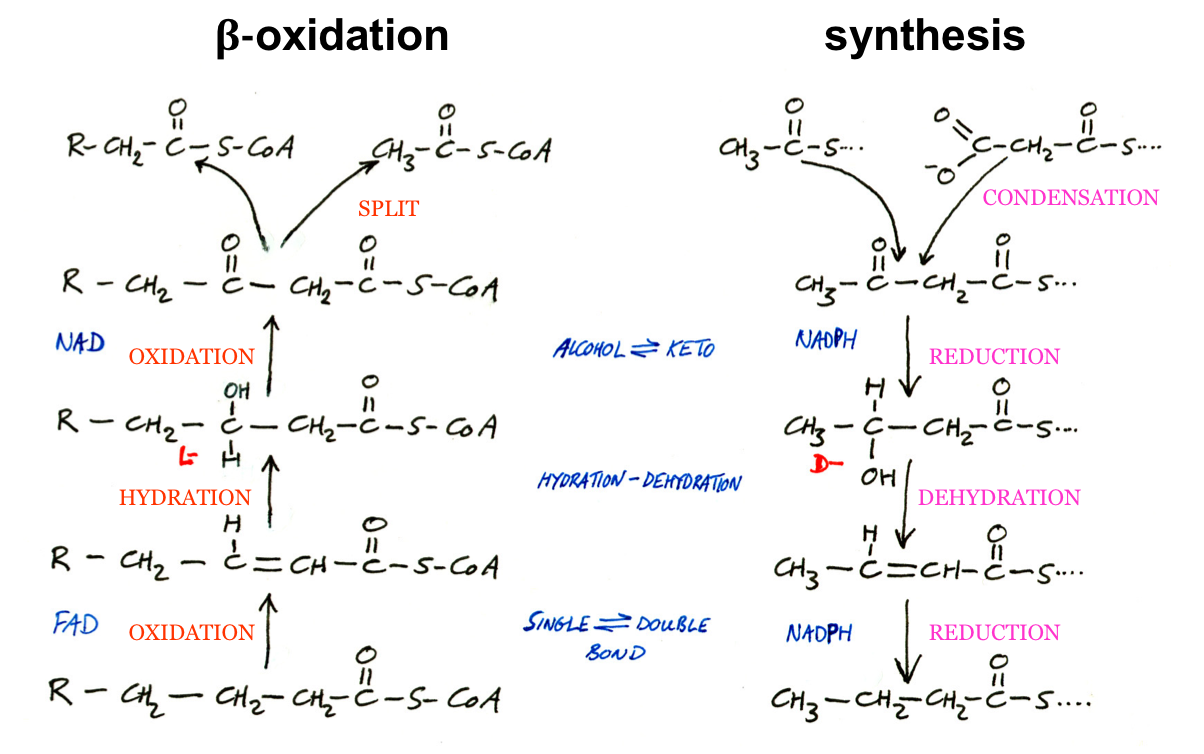
Acetyl-CoA is Transported from Mitochondria into the Cytosol as Citrate
Acetyl-CoA is made in the mitochondria.
Lipid synthesis occurs in the cytosol, but acetyl-CoA cannot cross the mitochondrial membrane.
Solution: Convert acetyl-CoA to citrate:
Acetyl-CoA + oxaloacetate → citrate (same as CAC reaction, catalyzed by citrate synthase).
Citrate crosses to the cytosol via the citrate transporter.
Citrate in Cytosol is Cleaved to Regenerate Acetyl-CoA
Citrate is cleaved by citrate lyase to regenerate:
Acetyl-CoA
Oxaloacetate
Reaction requires 1 ATP.
Acetyl-CoA can now be used to make malonyl-CoA.
Oxaloacetate is converted to Malate, which has two fates
Converted to malate by malate dehydrogenase.
Malate has 2 fates:
Converted to pyruvate + NADPH (via malic enzyme):
Pyruvate enters mitochondria and is converted to oxaloacetate.
NADPH is used for fatty acid synthesis.
Transported back to mitochondria as malate via malate-α-ketoglutarate transporter, then reoxidized to oxaloacetate.
SHUTTLE FOR TRANSFER OF ACETYL GROUPS FROM MITOCHONDRIA TO THE CYTOSOL
The mitochondrial outer membrane is freely permeable to all these compounds.
Pyruvate, derived from amino acid catabolism in the mitochondrial matrix or from glucose by glycolysis in the cytosol, is converted to acetyl-CoA in the matrix.
Acetyl groups pass out of the mitochondria as citrate; in the cytosol they are delivered as acetyl-CoA for fatty acid synthesis.
Oxaloacetate is reduced to malate, which can return to the mitochondrial matrix and be converted to oxaloacetate.
The major fate for cytosolic malate is oxidation by malic enzyme to generate cytosolic NADPH; the pyruvateproduced returns to the mitochondrial matrix.
ACETYL-CoAᶜʸᵗ AND BICARBONATE FORM MALONYL-CoA
Reaction carboxylates acetyl-CoA.
Catalyzed by Acetyl-CoA Carboxylase (ACC) (rate limiting step of fatty acid synthesis):
Enzyme has three subunits:
One unit has Biotin to carry CO₂.
HCO₃⁻ (bicarbonate) is the source of CO₂.
Uses one ATP molecule.
In animals, all three subunits are on one polypeptide chain.
Therefore, the cost of malonyl-CoA formation (precursor for fatty acid synthesis) is 3 ATPs per 2-carbon unit (2 for shuttling acetyl-CoA to the cytosol and 1 for making malonyl-CoA).
A REMINDER: BIOTIN CARRIES CO₂
Two-step reaction similar to carboxylations catalyzed by pyruvate carboxylase and propionyl-CoA carboxylase.
CO₂ binds to biotin:
Bicarbonate reacts with ATP to produce carboxyphosphate as an intermediate.
Carboxyphosphate breaks down to CO₂.
CO₂ is attached to N in ring of biotin.
Biotin transports CO₂ to acetyl-CoA on another active site.
THE ACETYL-CoA CARBOXYLASE (ACC) REACTION
Malonyl-CoA is formed from Acetyl-CoA and Bicarbonate
ACC has 3 functional regions:
Biotin carrier protein
Biotin carboxylase, which activates CO₂ by attaching it to a nitrogen in the biotin ring in an ATP-dependent reaction
Transcarboxylase, which transfers activated CO₂ (shaded green) from biotin to acetyl-CoA, producing malonyl-CoA.
The long, flexible biotin arm carries the activated CO₂ from the biotin carboxylase region to the transcarboxylase active site.
The active enzyme in each step is shaded blue in the diagram.
FATTY ACID SYNTHESIS OCCURS IN CELL COMPARTMENTS WHERE NADPH LEVELS ARE HIGH
Cytosol for animals (and yeast)
Major sources of NADPH:
In adipocytes: pentose phosphate pathway and malic enzyme:
NADPH made as malate converts to pyruvate + CO₂
In hepatocytes and mammary gland: pentose phosphate pathway
NADPH made as glucose-6-phosphate converts to ribulose-5-phosphate
To a lesser extent, NADP dependent isocitrate dehydrogenase reaction → isocitrate is oxidatively decarboxylated to α-ketoglutarate (depends on mitochondria)
SYNTHESIS OF FATTY ACIDS IS CATALYZED BY FATTY ACID SYNTHASE (FAS)
Catalyzes a repeating four-step sequence that elongates the fatty acyl chain by two carbons at each step.
Uses NADPH as the electron donor
Uses two enzyme-bound –SH groups as activating groups
FAS I in vertebrates:
Single polypeptide chain with several enzyme activities
Leads to single product: palmitate 16:0
C-15 and C-16 are from the acetyl CoA used to prime the reaction
FATTY ACID SYNTHESIS
Overall goal: attach two-carbon acetate unit from malonyl-CoA to a growing chain and then reduce it.
Reaction involves cycles of four enzyme-catalyzed steps:
Condensation of the growing chain with activated acetate
Reduction of carbonyl to hydroxyl
Dehydration of alcohol to trans-alkene
Reduction of alkene to alkane
The growing chain is initially attached to the enzyme via a thioester linkage.
During condensation, the growing chain is transferred to the acyl carrier protein (ACP)
After the second reduction step, the elongated chain is transferred back to fatty acid synthase.
ACYL CARRIER PROTEIN (ACP) SERVES AS A SHUTTLE IN FATTY ACID SYNTHESIS
Contains a covalently attached prosthetic group 4’-phosphopantetheine.
Flexible arm to tether acyl chain while carrying intermediates from one enzyme subunit to the next.
Delivers acetate (in the first step) or malonate (in all the next steps) to the fatty acid synthase.
Shuttles the growing chain from one active site to another during the four-step reaction.
FATTY ACID SYNTHASE TYPE I (FAS I)
Mammalian FATTY ACID SYNTHASE (FAS I): a mega synthase with multiple active sites
Condensation with acetate → β-ketoacyl-ACP synthase (KS)
Reduction of carbonyl to hydroxyl → β-ketoacyl-ACP reductase (KR)
Dehydration of alcohol to alkene → β-hydroxyacyl-ACP dehydratase (DH)
Reduction of alkene to alkane → enoyl-ACP reductase (ER)
Chain transfer/charging → Malonyl/acetyl-CoA ACP transferase (MAT)
Thioesterase → TE: releases the palmitate product from ACP
ACP → Acyl Carrier Protein
FATTY ACID SYNTHESIS PROCEEDS IN A REPEATING REACTION SEQUENCE
Addition of two carbons to a growing fatty acyl chain: a four-step sequence
Each malonyl group (on Acyl Carrier Protein; ACP) and acetyl (or longer acyl) group is activated by a thioester that links it to fatty acid synthase, then:
Condensation of an activated acyl group (an acetyl group from acetyl-CoA is the first acyl group) with two carbons derived from malonyl-CoA, with elimination of CO₂ from the malonyl group, extends the acyl chain by two carbon units.
Reduction of the β-keto group to an alcohol.
Dehydration (elimination of H₂O) → to create a double bond
Enzyme: DH (Dehydratase)
Reduction of the double bond → to create a saturated fatty acyl group
Enzyme: ER (Enoyl-ACP Reductase)
Translocation of the elongated product to an SH group on FAS → recharging the now free SH of ACP with another malonyl group
THE OVERALL PROCESS OF PALMITATE SYNTHESIS: SEVEN CYCLES OF CONDENSATION AND REDUCTION
The fatty acyl chain grows by two-carbon units donated by activated malonate, with loss of CO₂ at each step.
The initial acetyl group is shaded yellow, C-1 and C-2 of malonate are shaded pink, and the carbon released as CO₂ is shaded green.
After each two-carbon addition, reductions convert the growing chain to a saturated fatty acid of four, then six, then eight carbons, and so on.
The final product is palmitate (16:0).
STOICHIOMETRY OF SYNTHESIS OF PALMITATE (16:0)
Formation of 7 malonyl-CoA:
7 acetyl-CoAs are carboxylated to make 7 malonyl-CoAs… using ATP:7 Acetyl-CoA + 7 CO₂ + 7 ATP → 7 Malonyl-CoA + 7 ADP + 7 Pi7 Cycles of condensation, reduction, dehydration, and reduction:
Uses NADPH to reduce the β-keto group and trans-double bond:Acetyl-CoA + 7 Malonyl-CoA + 14 NADPH + 14 H⁺ → Palmitate + 7 CO₂ + 8 CoA + 14 NADP⁺ + 6 H₂OSum:
8 Acetyl-CoA + 7 ATP + 14 NADPH + 14 H⁺ → Palmitate + 8 CoA + 6 H₂O + 7 ADP + 7 Pi + 14 NADP⁺
Note: The total cost of fatty acid synthesis is 3 ATPs per 2-carbon unit
(2 for shuttling acetyl-CoA to cytosol, and 1 for making malonyl-CoA) → Minus 1 ATP for the first acetyl-CoA (not from malonyl-CoA).
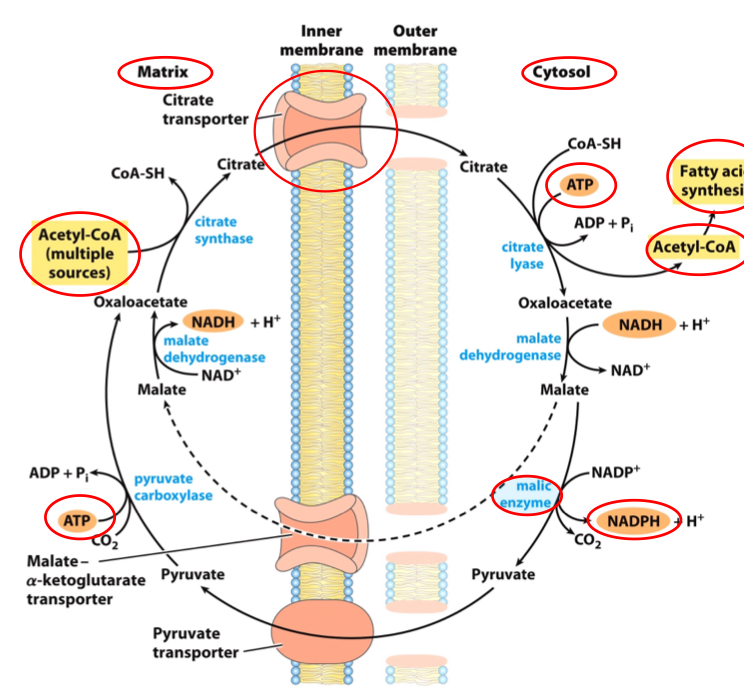
FATTY ACID SYNTHESIS IS TIGHTLY CONTROLLED VIA ALLOSTERIC REGULATION OF ACC
Acetyl-CoA Carboxylase (ACC) catalyzes the rate-limiting step
Inhibited by palmitoyl-CoA
Activated by citrate
Citrate is made from acetyl-CoA
When [acetyl-CoA]⁽ᵐᵗ⁾ ↑ (high ATP), citrate is exported → activates ACC
Citrate signals excess energy to be converted to fat
Note: Citrate also inhibits glycolysis by inhibiting PFK-1
ACC IS ALSO HORMONALLY REGULATED BY COVALENT MODIFICATION
ACC is inhibited when energy is needed (low glucose):
Glucagon and epinephrine:
↓ Citrate activation sensitivity
→ Phosphorylation & inactivation of ACC
(Phosphorylated monomers of ACC are inactive)
ACC is activated when energy is plenty (high glucose):
Insulin:
→ Dephosphorylation of ACC monomers
→ Polymerization into long active ACC filaments
RECAP: REGULATION OF FATTY ACID SYNTHESIS IN VERTEBRATES
Regulation of fatty acid synthesis:
(a) In vertebrates, allosteric & hormone-dependent covalent regulation influence precursor flow to malonyl-CoA
(b) ACC filaments (active, dephosphorylated form) are observed via electron microscopy
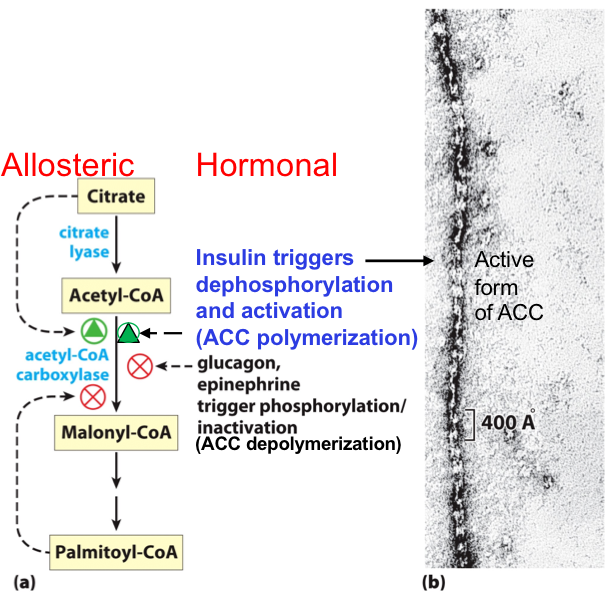
COORDINATED REGULATION OF FATTY ACID SYNTHESIS AND BREAKDOWN
Two enzymes are key to the coordination of fatty acid metabolism: acetyl-CoA carboxylase (ACC) (the first enzyme in the synthesis of fatty acids), and carnitine acyltransferase I (which limits the transport of fatty acids into the mitochondrial matrix for β-oxidation).
Synthesis: Acetyl-CoA Carboxylase (ACC)
Oxidation: Carnitine Acyl Transferase I (CAT I)
REGULATION TARGETS
When the diet provides a ready source of carbohydrate as fuel, β-oxidation of fatty acids is unnecessary and is therefore downregulated.
Ingestion of a high-carbohydrate meal raises the blood glucose level and thus triggers the release of insulin.
Insulin triggers the release of protein phosphatase which dephosphorylates ACC, activating it.
ACC catalyzes the formation of malonyl-CoA (the first intermediate of fatty acid synthesis).
Malonyl-CoA inhibits carnitine acyltransferase I, thereby preventing fatty acid entry into the mitochondria.
When blood glucose levels drop between meals:
Glucagon is released and activates cAMP-dependent protein kinase (PKA).
PKA phosphorylates and inactivates ACC.
The concentration of malonyl-CoA falls, the inhibition of fatty acid entry into mitochondria is relieved.
Fatty acids enter the mitochondrial matrix.
β-oxidation becomes the major fuel source.
Because glucagon also triggers the mobilization of fatty acids in adipose tissue, a supply of fatty acids begins arriving in the blood.
Glucagon also triggers mobilization of fatty acids from lipid droplets in adipose tissue (through phosphorylation of HSL).
Also, transcriptional regulation of CAT, acyl CoA dehydrogenases, etc., contributes to long-term adaptation.
LONGER-CHAIN AND UNSATURATED FATTY ACIDS ARE SYNTHESIZED FROM PALMITATE
Routes of synthesis of other fatty acids:
Elongation of Palmitate (16:0) → Stearate (18:0)
Catalyzed by elongase
Adds two carbons at COOH end
Uses malonyl-CoA, just like fatty acid synthase (FAS)
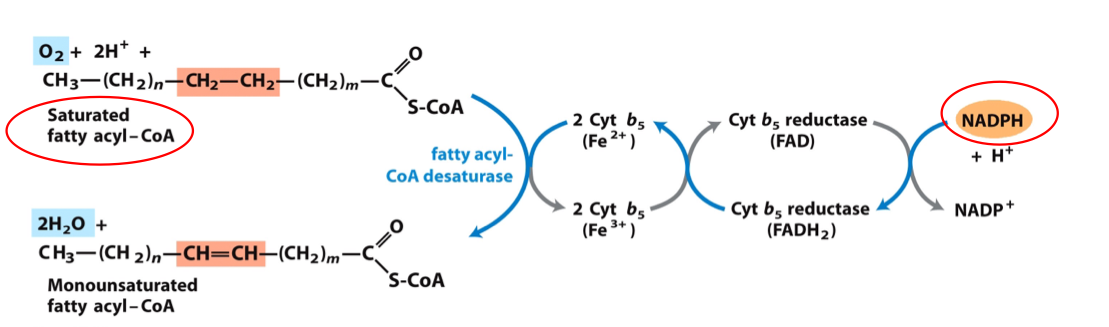
Desaturation of Palmitate (16:0) → Palmitoleate (16:1Δ9)
Stearate (18:0) → Oleate (18:1Δ9)
Catalyzed by fatty acyl-CoA desaturase
Requires NADPH
Uses cytochrome b₅ and cytochrome b₅ reductase
This is a Δ9-desaturase! It reduces the bond between C-9 and C-10
Mammals cannot desaturate beyond Δ9 → cannot make linoleate or α-linolenate
➝ Must be obtained in diet → essential fatty acidsConversion of α-linolenate (ALA) to:
EPA: 20:5 (Δ5,8,11,14,17)
DHA: 22:6 (Δ4,7,10,13,16,19)
Requires elongation/desaturation
PLANTS, BUT NOT MAMMALS, CAN DESATURATE POSITIONS BEYOND C-9
Humans have Δ4, Δ5, Δ6, and Δ9 desaturases
But cannot desaturate beyond Δ9
Plants can produce:
Linoleate (18:2, Δ9,12)
α-Linolenate (18:3, Δ9,12,15)
These are essential fatty acids for humans and are precursors to:
PUFAs (polyunsaturated fatty acids) → membrane fluidity, precursors to eicosanoids
VERTEBRATE FATTY ACYL DESATURASE: NON-HEME IRON-CONTAINING OXIDASE
O₂ accepts 4 electrons total:
2 from saturated fatty acid
2 from ferrous state of cytochrome b₅ / NADPH
Located in smooth ER
Electrons flow through:
Fatty acyl-CoA → NADPH → desaturation pathway
BIOSYNTHESIS OF TRIACYLGLYCEROLS
Phosphatidic acid is the precursor for both TAGs and glycerophospholipids
Lipin (phosphatidic acid phosphatase) removes the phosphate:
Yields 1,2-diacylglycerol
Third carbon is acylated with another fatty acid:
Forms triacylglycerol (TAG)
BIOSYNTHESIS OF MEMBRANE LIPIDS
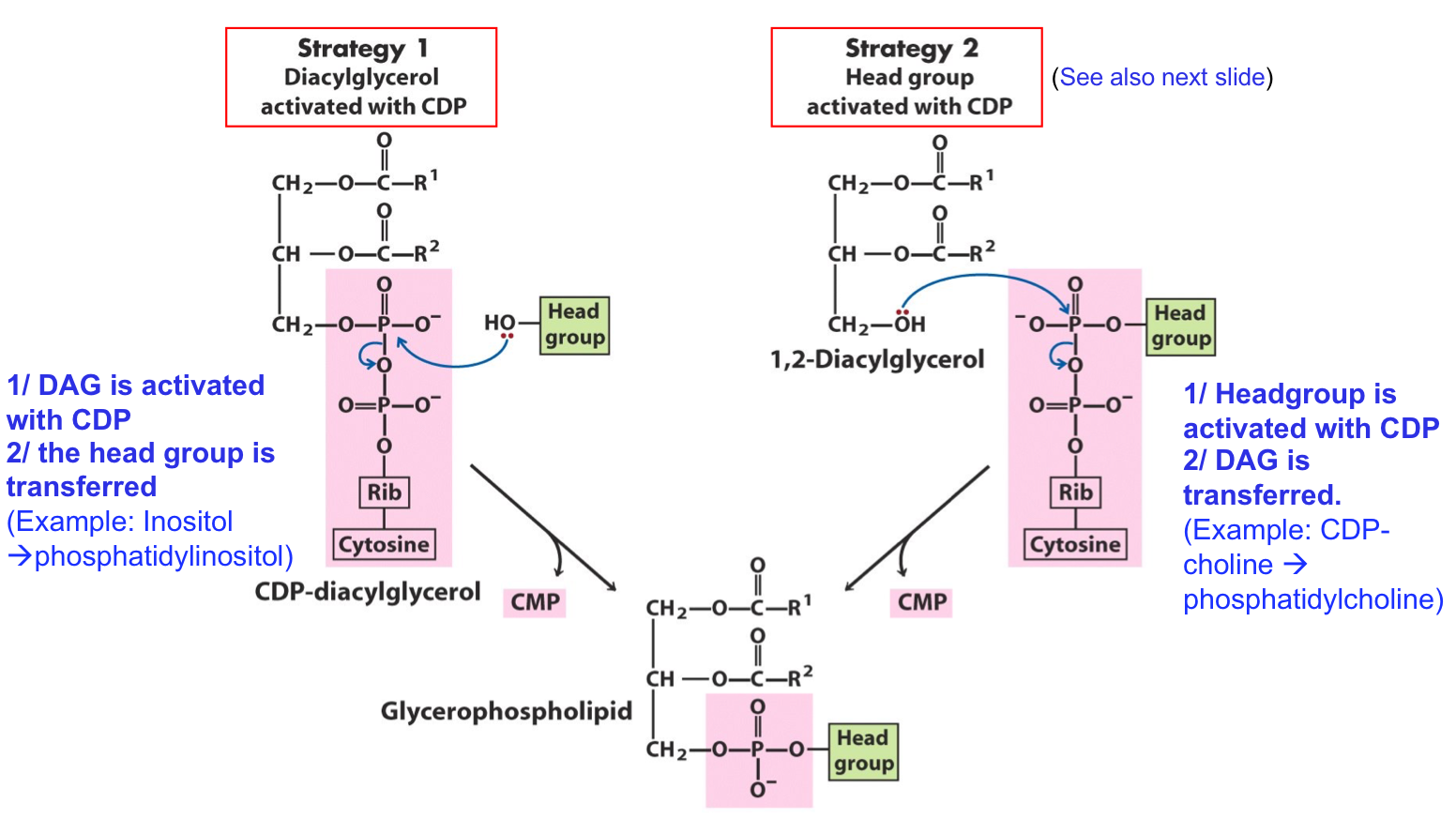
PHOSPHODIESTER BOND OF PHOSPHOLIPIDS FROM CDP
Two general strategies for forming the phosphodiester bond of phospholipids:
In both cases, CDP supplies the phosphate group of the phosphodiester bond.
Strategy 1: Diacylglycerol activated with CDP
DAG is activated with CDP
The head group is transferred
Example: Inositol → phosphatidylinositol
Strategy 2: Head group activated with CDP
Headgroup is activated with CDP
DAG is transferred
Example: CDP-choline → phosphatidylcholine
BIOSYNTHESIS OF MEMBRANE LIPIDS
SPHINGOLIPIDS ARE MADE IN FOUR STEPS
Synthesis of sphinganine from palmitoyl-CoA and serine
Attachment of fatty acid via amide linkage
Desaturation of sphinganine
Yields N-acylsphingosine (ceramide)
Attachment of head group
Can yield a cerebroside or ganglioside
Summary of Fatty Acid Biosynthesis and Degradation
Fatty acid synthesis and degradation consist of cycles to add or remove 2-carbon units, governed by distinct but coordinated pathways.
Key regulatory enzymes involved include Acetyl-CoA Carboxylase and Carnitine Acyltransferase I.