Cell Junctions and the Extracellular Matrix 04-15-2025
Cell Junctions:
Cell junctions are little adhesions that form between cells
Fundamental for the structural integrity and organization of tissues in multicellular organisms.
Hold cells together through various types of junctions, ensuring they function as cohesive units.
Maintain multicellular "tissue" organization by allowing for specialized functions within different tissue types.
Metazoan Organisms: Met-a-zon”—>Multi-animal-cells”
Functions:
(1) Hold two adjacent cells together
Hold two adjacent cells together If we have two adjacent cells and specific proteins that were anchored between two cells to hold them together
If there are any kinds of shearing forces, stretching, or abrasive forces, → Resist the separation of the cells
Hinder abrasive, shearing, and stretching forces, it can withstand those forces
(2) Diffusion barrier
If you don’t want Ions to move through the cell, and we don’t want certain things to move through, it will block.
Tight junctions can block them → act like a diffusion barrier
Facilitating the movement of materials (e.g., ions, small molecules) between joined cells, which is crucial in tissues like heart muscle, where cell-to-cell signaling influences contraction.
(3) Allow cell communication
Establishing communication pathways that allow cells to exchange signals and materials vital for maintaining homeostasis.
Little channels that exist between the two cells
Allows cell communication
(4)Connects cell to the basal lamina
Structure under the cells → basal lamina / extracellular matrix (ECM)
Basal lamina is a little bit of connective tissue.
Besides connecting cell-to-cell, adhesion molecules also connect cells to the basal lamina.
This makes up the basement membrane
Extracellular Matrix (ECM):
The ECM is a complex network composed of proteins, polysaccharides, and other macromolecules, providing structural and biochemical support to surrounding cells.
Synthesized and secreted by tissue cells create a unique environment that supports cellular functions and tissue integrity.
Closely associated with producing cells, allowing dynamic responses to physiological changes.
Most elaborated in connective tissues, where ECM is more plentiful than the cells it supports, defining the tissue's properties.
Exhibits varied forms due to the compositional and organizational differences of macromolecules, which are adapted to specific tissue requirements.
Methods of Cohesion
Direct cell-to-cell interactions via junctions.
Extracellular Matrix (ECM): secreted by cells; network of proteins & polysaccharides.
Importance of Cell Cohesion
Essential for forming organized multicellular structures.
Allows tissues to withstand external forces.
Governs body architecture: shape, strength, cell arrangement.
Assembled locally by cells into a complex, organized mesh.
Found outside cells but tightly integrated with them.
It is where the basal lamina resides
Who Builds the ECM?
Cells living in the matrix make and organize it
Fibroblasts: Main ECM producers in connective tissue
Chondroblasts: Make cartilage ECM
Osteoblasts:Make bone ECM
Epithelial: all these cells are primarily responsible for producing the various components of the extracellular matrix.
What is it made of?
GAGs (Glycosaminoglycans),Fibrous Proteins,Non-collagen Glycoproteins
Roles of Cell Attachments
Enable cell movement during growth, development, and repair.
Influence cell orientation and behavior via the cytoskeleton.
Allow cells to sense/respond to mechanical changes in the environment.
Matrix Molecules:
Hyaluronan, Proteoglycans, Fibronectin, Laminin—are part of the extracellular matrix ensemble
Glycosaminoglycan (GAG):
Comprised of unbranched negativly polysaccharide chains of repeating disaccharides that contribute to the gel-like properties of the ECM.
A highly negative charge creates a gel at low concentrations, which is vital for hydration and nutrient diffusion.
Composition
Resist compression + fill space
Long/Extended, stiff/rigid → take up lots of space
Do not fold like proteins
ECM fillers: Even though they're light in weight (<10% ECM protein weight), they fill huge amounts of space!
Classification based on
type of linkage between sugars
number/position of sulfate groups
Function:
Attract water → gel-like consistency
Large (-) charge attracts cations water follows osmotically
Very hydrophilic = sucks in water like a sponge
Usually attached to proteins → become proteoglycans
Compression resistance: All that water creates turgor pressure, making tissues shock-absorbent (ex: knee cartilage under pressure).
Defects = Big Trouble: A lack of dermatan sulfate → developmental issues: short stature, early aging, skin/joint/muscle problems.
Hyaluronan:
The simplest type of GAG consists of non-sulfated disaccharides that play a critical role as a "space-filler" in tissues and are energetically inexpensive to produce.
Simple Structure: Unlike other GAGs that have different sugar repeats, hyaluronan has a single repeating disaccharide unit, which makes it simpler and more straightforward.
No Sulfation: Other GAGs usually have sulfate groups, but hyaluronan doesn’t, which is another reason it's unique.
Huge Size: Hyaluronan can be massive, with chains up to 25,000 disaccharide units long, giving it a huge surface area and making it important for filling space and creating volume in tissues.
No Core Protein: While most GAGs are linked to proteins (to form proteoglycans), hyaluronan is free-floating and not typically bound to a core protein.
Synthesis: While most GAGs are synthesized inside the cell and then secreted, hyaluronan is made at the cell surface through an enzyme complex embedded in the plasma membrane. This gives it a very different synthesis route!
Functions of Hyaluronan:
Compression Resistance: It’s great at resisting compressive forces in tissues, making it important for joint fluid (synovial fluid) and cartilage.
Developmental Role: Hyaluronan helps shape tissues during embryonic development by expanding with water and forcing tissue structures to change shape. For example, it helps the formation of the heart valves and septa.
Wound Healing & Lubrication: During wound healing, hyaluronan is produced in large amounts. It's also found in joint fluid, providing lubrication to prevent friction between bones.
Proteoglycans:
Formed from GAGs covalently attached to core proteins, they regulate the activity of various secreted molecules and can bind specific signal molecules, acting as co-receptors when associated with cell surfaces.
Core Protein & GAGs: Unlike hyaluronan, which floats around on its own, proteoglycans are made of a core protein that gets covalently attached to glycosaminoglycan (GAG) chains. This gives them a heavy carbohydrate load (about 95% carbohydrate by weight) and a long, unbranched structure.
Synthesis: These core proteins are first synthesized by ribosomes and enter the endoplasmic reticulum. GAGs are then attached to the core protein in the Golgi apparatus—this process includes adding a special linkage tetrasaccharide to a serine side chain to kickstart the GAG addition.
Regulate activity of secreted molecules selective sieves regulate intercellular traffic, bind secreted signal molecules and influenctheir behavior, and some proteoglycans associate with the cell surface and function as co-receptor
GAG Chain Diversity: GAG chains can be modified with sulfate groups to increase negative charge, and the repeating sugar units can undergo changes that give proteoglycans lots of structural diversity.
Proteoglycan Size: Some proteoglycans, like aggrecan, are gigantic, with over 100 GAG chains! Others, like decorin, are much smaller, having only one GAG chain. Huge diversity of carbohydrate and protein components, these sizes give them different roles in the matrix.
Functional Roles of Proteoglycans:
Tensile Strength & Matrix Assembly: Proteoglycans like decorin are involved in regulating collagen fibril assembly, which affects the tensile strength of tissues like skin. A lack of decorin in mice leads to fragile skin—clearly, these guys are key players!
Aggregates & Complexes: Proteoglycans can form massive aggregates. For instance, aggrecan in cartilage combines with hyaluronan to form large, bacterial-sized complexes that help resist compressive forces in joints. This assembly gives cartilage its shock-absorbing properties.
Cell Signaling: Some proteoglycans aren't just matrix components—they're also involved in cell signaling. Syndecans, for example, are membrane-bound proteoglycans that interact with the actin cytoskeleton and signaling proteins inside cells, helping regulate cell behavior.
Membrane-Bound Proteoglycans (Syndecans):
Syndecans are a special class of proteoglycans that are embedded in the plasma membrane. These proteoglycans span the membrane and interact with the cytoskeleton, regulating both mechanical properties of the cell surface and signaling events.
Anchored to the Membrane: Some syndecans have glycosylphosphatidylinositol (GPI) anchors that tie them to the lipid bilayer, keeping them connected to the cell surface.
Examples include
Chondroitin sulfate, Heparan sulfate, and Keratin sulfate, which are classified based on sugar linkage and sulfate groups.
Decorin, perlecan (protein + GAGs) which help fill space, resist compression, bind growth factors.
Fibrous Proteins:
Collagen:
Collagens are the heavyweights of the extracellular matrix, making up a huge portion of the protein mass in mammals—around 25%. They are a family of fibrous proteins that are present in nearly all multicellular animals, playing essential roles in the structural integrity of tissues. Let's dive into the fascinating world of collagens:
triple-Helical Structure: The defining feature of collagen is its long, stiff, triple-stranded helical structure. Imagine three α chains (polypeptide chains) twisted around each other like a rope in a superhelix. This highly organized structure is essential for the strength and rigidity of collagen fibers.The main protein in ECM that forms a stiff structure composed of triple-stranded polypeptide chains, providing tensile strength through covalent and ionic bonds.Thus Provide structure + strength.
Amino Acid Composition: Collagen is rich in proline and glycine, both of which are crucial for the stability of the triple helix. Glycine, being the smallest amino acid, fits perfectly into the center of the helix, allowing the strands to coil tightly.
Network-Forming Collagens: Type IV collagen forms a mesh-like network and is found in the basal lamina, a thin layer of extracellular matrix that underlies epithelial cells.
Anchoring Fibrils: Type VII forms dimeric structures that help anchor the basal lamina to the underlying tissue. These are particularly important in tissues like skin.
Elastin:
Elastic tissues, such as those in the skin, blood vessels, and lungs, have a critical need for both strength and elasticity—meaning they must be able to stretch and return to their original shape without tearing
Structure & Function: Elastic fibers are extensible—they can stretch up to five times their resting length. This is far more than the stretchability of a rubber band of the same size. Collagen fibrils, which are inelastic, are interwoven with elastic fibers, limiting the stretch and preventing tearing. Together, they balance strength and flexibility.
The Core Protein: Elastin is the main protein in elastic fibers. Unlike collagen, elastin is highly hydrophobic and rich in proline and glycine. However, unlike collagen, elastin is not glycosylated, meaning it doesn’t have sugar chains attached to it. It consists of about 750 amino acids and is produced in a precursor form called tropoelastin.
Cross-Linking: Tropoelastin is secreted into the extracellular matrix, where it undergoes cross-linking with other tropoelastin molecules to form the elastin fibers. This cross-linking process is essential to give elastin its rubber-like properties—it’s what allows the material to stretch and recoil.
Composition:
Hydrophobic and Alpha-Helical Segments: The elastin protein is made up of alternating segments:
Hydrophobic segments: Responsible for the elastic properties of elastin, allowing it to stretch and return to shape.
Alanine- and lysine-rich α-helical segments: These segments form cross-links between elastin molecules, helping form the strong network that gives the fiber its strength and stability.
Conformation: Elastin’s structure is still somewhat mysterious, but it's thought that parts of the protein adopt a random coil conformation, much like regular rubber polymers. This random coil configuration is key to the fiber’s ability to stretch and recoil when needed (just like a rubber band).
Elastin Aorta
Elastin is a dominant extracellular matrix protein in the aorta—it makes up about 50% of the dry weight of the largest artery in the body. This elasticity is crucial for arteries to maintain blood pressure and prevent damage due to pressure changes. Mutations leading to elastin deficiencies can cause arteries to lose their elasticity, leading to conditions like narrowing of the aorta and excessive smooth muscle cell proliferation in the arterial wall. Without the proper elasticity, blood vessels lose their resilience and can become prone to rupture.
Non-collagen Glycoproteins
Fibronectin:
An adhesive protein that links matrix proteins to cell surface proteins, facilitating cell adhesion and migration.
Synthesis of Peptides: One way to study fibronectin's function is by synthesizing individual regions of the protein and testing their ability to bind to other molecules. For example, fibronectin can bind to:
Collagen
Proteoglycans
Integrins on the surface of various types of cells.
Laminin:
Laminins comprise a large family of proteins, each composed of three long polypeptide chains (α, β, and γ) held together by disulfide bonds and arranged in the shape of an asym- metric bouquet, like a bunch of three flowers whose stems are twisted together at the foot but whose heads remain separate
A key glycoprotein that provides a scaffold for cell adhesion, influencing cell differentiation and shape. Glue-like, help with adhesion & signaling
Cellular Roles:
Determines Cell Polarity: It helps establish the orientation of cells, ensuring proper function and organization.
Influences Cell Metabolism: The basal lamina can regulate cellular activities, affecting metabolism and function.
Organizes Plasma Membranes: It plays a role in organizing proteins in the adjacent plasma membranes, ensuring correct cell-cell and cell-matrix interactions.
Promotes Cell Survival, Proliferation, and Differentiation: By influencing the environment of cells, the basal lamina can help maintain cell health and function, guiding growth and differentiation.
Cell Migration: The basal lamina provides a scaffold or "highway" for cell movement, which is important during processes like embryogenesis, wound healing, and immune response.
Cell adhesion
Signaling
Organizing ECM architecture
Tissues:
Structures within an organism are relatively large and sturdy, allowing them to withstand various stresses and forces.
Formed from small, squishy structures with flimsy plasma membranes, which makes cellular attachment mechanisms vital.
It involves intricate connections between cytoskeletons of adjacent cells and interactions with an extracellular matrix, which provides mechanical support.
Attachment modifications are essential over time, adapting to mechanical demands and developmental cues.
Main Tissue Categories:
Connective Tissue:
Form: bone or tendon, which are formed from an extracellular matrix produced by cells that are distributed sparsely in the matrix.
Characterized by a high matrix-to-cell ratio, providing structural support across various organs.
Most mechanical stress is borne by fibrous matrix proteins, allowing it to withstand tension and compression.
Cell-cell adhesions are rare; their primary role is to support and bind other tissues. Direct attachments between one cell and another are relatively rare, but the cells have important attachments to the matrix.
Function
Sparse cell distribution.
ECM bears the most mechanical stress.
Few direct cell-cell connections.
Strong cell-matrix attachments link the cytoskeleton to ECM.
Epithelial Tissue:
Contains sparse ECM, primarily consisting of basal lamina, which is crucial for providing a substrate for cell adhesion.
Transmembrane proteins mediate cell adhesion and bind to the basal lamina, facilitating tissue integrity and function.
Most mechanical stress is managed by cell junctions, reinforcing structural connections between epithelial cells.
Structure of Epithelial Tissue
Found in surfaces like skin and the gut lining.
Cells form tightly packed sheets called epithelia.
ECM is minimal—mainly a thin layer called the basal lamina (or basement membrane).
Cells are connected by direct cell–cell junctions.
The cytoskeleton of each cell transmits mechanical stress through these junctions.
Image

Eliphtical Junctions
Epithelial sheets line surfaces, cavities, and internal compartments of the animal body.
Thought to be a key evolutionary innovation in animals.
Epithelial cells show structural polarity:
Basal side: attached to other tissues via basal lamina
Apical side: exposed to extracellular fluid
Selective permeability barrier between basal and apical fluids.
Requires tight junctions to:
Prevent leakage of molecules between cells
Maintain chemical differences between apical and basal environments


Anchoring Junctions
Connect cells through the cytoskeleton to neighboring cells or the ECM, preserving tissue architecture.
Include transmembrane proteins such as cadherins (cell-to-cell connections) and integrins (connecting cells to the basal lamina).
Symmetry in Anchoring Junctions
Junctions between cells are typically symmetrical:
Actin in one cell ↔ Actin in the neighboring cell.
Transmembrane Adhesion Proteins
transmembrane adhesion proteins that span the plasma membrane, with one end linking to the cytoskeleton inside the cell and the other end linking to other structures outside transmembrane adhesion proteins
Link:
Inside: to the cytoskeleton.
Outside: to other cells (via cadherins) or ECM (via integrins).
Form the core of anchoring junctions.
All JAMs are transmembrane adhesion proteins, but not all transmembrane adhesion proteins are JAM.

Cadherin & Integrins
Cadherin Family (Cell-Cell Adhesion):
1. What Are Cadherins?
Calcium-dependent cell–cell adhesion proteins.
Name = Ca²-dependent adhesion → “cadherin.”
Removing Ca²⁺ = adhesion falls apart.
Essential for anchoring junctions like:
Link to actin → form Adherens Junctions
Link to intermediate filaments → form Desmosomes
Two or more cadherins associate to form oligomers.
2. Structure & Binding
Cadherins have multiple EC (extracellular cadherin) domains.
Binding site: at the N-terminal tips (farthest from the membrane).
One cadherin's "knob" fits into the other’s "pocket".
Ca²⁺ ions bind at hinge regions between EC domains:
Calcium keeps cadherins rigid.
Ca²⁺ binds at hinge regions between cadherin domains.
This prevents flexing, maintaining a stiff rod-like structure.
Make the cadherin rigid and curved like a springy rod
Without Ca²⁺:
Hinge regions flex.
The structure becomes floppy and unstable.
Weakens N-terminal binding affinity, leading to disassembly of cell–cell adhesion.
Without Ca²⁺, cadherins become floppy, reducing adhesion.
Cadherin Binding (Outside):
Extracellular domains mediate homophilic binding between cadherins on adjacent cells.
Stabilizes cell–cell adhesion.
Cadherin Binding (Inside):
Intracellular domains of cadherins connect to the cytoskeleton, but indirectly via adaptor proteins.
Cadherin binding is homophilic:
Binds to the same or closely related cadherin subtype on adjacent cells.
They are not general-purpose glue; they’re selective matchmakers.
Different cell types express different cadherins → promotes self-recognition and segregation from others. This specificity helps maintain tissue integrity and organizes cells into distinct structures.
3.Location
Attachments typically involve homophilic, homologous symmetrical associations, where similar cadherins engage with one another.
4. Binding Strength: The Velcro Principle
Individual cadherin interactions = low affinity.
Strength comes from numbers: many cadherins binding side-by-side in clusters.
Like Velcro: many small weak hooks = one strong hold.
This design allows:
Strong adhesion overall.
Easy disassembly when needed (e.g., during tissue remodeling).
Weak individually, strong together = adaptable but sturdy structure.
5. Importance
Cadherins without their cytoplasmic domains → no stable adhesion.
6. Evolutionary Importance
Found in all multicellular animals analyzed.
Also present in choanoflagellates (unicellular/multicellular protists).
Absent from:
Plants Fungi Bacteria Archaea
Suggests cadherins are a defining feature of animals.
Integrin Family (Cell-Matrix Adhesion):
Heterodimeric proteins: made of one α (alpha) and one β (beta) subunit.
Each subunit is transmembrane with a large extracellular domain.
Different combinations of α and β give rise to many integrin types, each with specific ligand-binding preferences.
Bind to ligands, such as fibronectin, collagen, and laminin, facilitating cell adhesion and communication within the extracellular matrix.
Not static, its able to change in response to different environmental signals, allowing tissues to adapt and communicate during development and repair processes.
They mediate important information with the use of the basal lam.
Link to actin → form Actin-linked Cell–Matrix Junctions
Link to intermediate filaments → form Hemidesmosomes
Integrin Activation:
Outside In
1) matrix protein activates integrin
2) integrin binds talin
3) talin binds actin
4) new cell shape results
Inside Out
1) talin is phosphorylated
2) talin bridges actin and integrin
3) integrin adopts new conformation
4) integrin “grabs” matrix
Feature
📌 Function
Anchoring cells to ECM
Mechanical linkage between ECM and cytoskeleton (especially actin filaments)
Involved in:
Cell migration
Wound healing
Immune response
Tissue structure maintenance
Integrins
Cell-ECM adhesion + signal relay
Low affinity, high abundance
Velcro-style binding
Low-affinity binding, but high surface density → strong adhesion via Velcro principle
Bidirectional signaling:
Outside-in: ECM → integrin →/signaling pathways
Inside-out: intracellular signals → integrin conformation change → ECM binding
Focal adhesions
Integrin clusters + cytoskeletal & signaling proteins
Anchorage dependence
ECM contact = life; no contact = apoptosis
Cell survival, proliferation, and growth require ECM attachment
Detachment → apoptosis
FAK, Talin, ILK
Signaling + scaffolding pros
Cancer cells
Often escape anchorage control
Cancer cells often override anchorage dependence, aiding invasion and survival in abnormal environments
Tight Junctions:
Definition: Specialized connections between adjacent epithelial cells that form a barrier to prevent the passage of molecules and ions through the space between cells. They are considered cell-cell junctions that play a critical role in maintaining the integrity of epithelial layers.
How: Tight junctions hold the cells closely together near the apex, sealing the gap between the cells and thereby preventing molecules from leaking across the epithelium
Freeze-fracture EM reveals a branching network of sealing strands that fully encircle the apical end of each epithelial cell.
Sealing strands: tightly apposed outer leaflets of neighboring plasma membranes.
Each strand = rows of transmembrane adhesion proteins with homophilic interactions (same protein binding to same protein across membranes).
These seal the intercellular space, blocking diffusion between cells.
Lumen
↓
[ Apical surface ]
[ Cell body ]
[ Basal surface ] ← basement membrane
Location: Found in epithelial and endothelial tissues, especially in areas like the intestines, kidneys, and blood-brain barrier.
Structure:
Composed of transmembrane proteins such as:
Claudins
Primary structural proteins of tight junctions.
Essential for tight junction formation and barrier function.
Claudins and occludins are two particular proteins that span out from the cell membrane into the ECM and they connect with one another. They interact with one another in the space between the cells.
Anchoring two cells to form tight junctions, which are essential for maintaining the polarity of cells and regulating the passage of substances between them.
Can form paracellular pores for selective ion transport (e.g., Mg²⁺ in kidneys).
Different claudins = different ion permeability profiles.
Kidney example: a claudin is required to resorb Mg²⁺ via the paracellular route.
Mutation in that claudin gene → Mg²⁺ loss in urine.
Occludins
Prevent the diffusion of substances between cells and maintain membrane integrity.
Composed of claudin and occludin proteins, forming tight junctions, which are crucial in maintaining selective permeability.
Claudins and occludins interact extracellularly like cadherins do in adherens junctions → promotes tight junction assembly.
Help maintain barrier function, especially in response to stress or signaling cues.
Intracellular organization relies on scaffold proteins binding the cytoplasmic tails of claudins/occludins.
Vertebrates—tight junctions
Invertebrates—septate junctions
Zona occludins(scaffold proteins)
Proteins that are located on the cytosolic side
Different types of zona occludens
ZO-1
ZO-2
ZO-3
These actins attach themselves to the Zona or are linked to the actin cytoskeleton via scaffold proteins like ZO-1, ZO-2, and ZO-3.
Multiple PDZ domains (bind to C-terminal tails of partner proteins).
Ability to bind to other ZO proteins → forms an intracellular protein mat that stabilizes sealing strands.
Junctional adhesion molecules (JAMs)
belong to the immunoglobulin superfamily.
Found primarily at tight junctions in epithelial and endothelial cells.
Have a single transmembrane domain.
Enable homophilic and heterophilic interactions via their extracellular regions.
Connect to the cytoskeleton through scaffold proteins like ZO-1.
Purpose:
Help form and maintain tight junctions.
Support cell polarity and barrier integrity.
Facilitate leukocyte transmigration in immune responses.
Participate in cell signaling.
Function
Strength It holds the cells tightly together, thereby providing structural integrity to tissues and regulating the flow of materials between cells.
The Basal Lamina:
A thin, flexible layer of ECM acts as a selective filter between cells and connective tissues, playing a crucial role in tissue organization, function, and mechanical state.
It’s found in various locations throughout the body, including beneath epithelial cells and between underlying connective tissues, i.e, surrounding muscle cells, thus providing support and anchorage to these tissues.
Purpose:
Acts as a supportive scaffold for cells.
Anchors epithelial cells via hemidesmosomes.
Regulates cell behavior, including differentiation, migration, and survival.
Serves as a selective filter (e.g., in the kidney glomerulus).
Helps in tissue regeneration and compartmentalization.
Apical Surface:
The apical surface is the topmost part of an epithelial cell that faces outward—toward the lumen (internal space) or the external environment.
They help Tight junctions connect cells to cells at the apical surfaces.
Which play a crucial role in regulating the passage of molecules between adjacent cells, ensuring that substances reach their intended destinations, and maintaining the integrity of the tissue.
Purpose:
Absorption (e.g., nutrients in the intestine)
Secretion (e.g., mucus in respiratory tract)
Protection from physical or chemical damage
Sensation in specialized tissues
Together:
Function as molecular “fences” that prevent mixing of membrane proteins between:
Apical membrane (facing lumen)
Basolateral membrane (facing underlying tissue)
Diffusion barrier:
A diffusion barrier is a physical or biochemical structure that restricts or controls the movement of molecules (especially solutes and water) between compartments.
In tight junctions, they contribute to signal transduction, allowing cells to communicate effectively while maintaining their polarized structure.
Maintains selective permeability—only certain substances can pass through, thus preserving the distinct environments necessary for proper cell function (homeostasis).
EX: Imagine Na+ or K+ ion or protein molecule that we don’t want them to be able to move between the cells
They act as diffusion barrier
Block the transport of ions and different types of large molecules between the cells
Helps establish concentration gradients by preventing the free passage of ions and other small molecules, which is essential for cellular function and signaling.
Role:
Protects internal environments from harmful substances.
Regulates nutrient uptake and waste removal.
Maintains homeostasis by controlling molecular movement.
Supports cell polarity and organized tissue function.
Seal neighboring cells together to block paracellular transport
Functions:
Barrier function: Regulates paracellular transport (movement between cells).
Fence function: Maintains cell polarity by separating apical and basolateral membrane domains.
Signal regulation: Involved in cell signaling pathways.
Images



Clinical Ref.
Clinical Relevance:
Dysfunction is linked to diseases like inflammatory bowel disease, leaky gut, and neurological disorders.

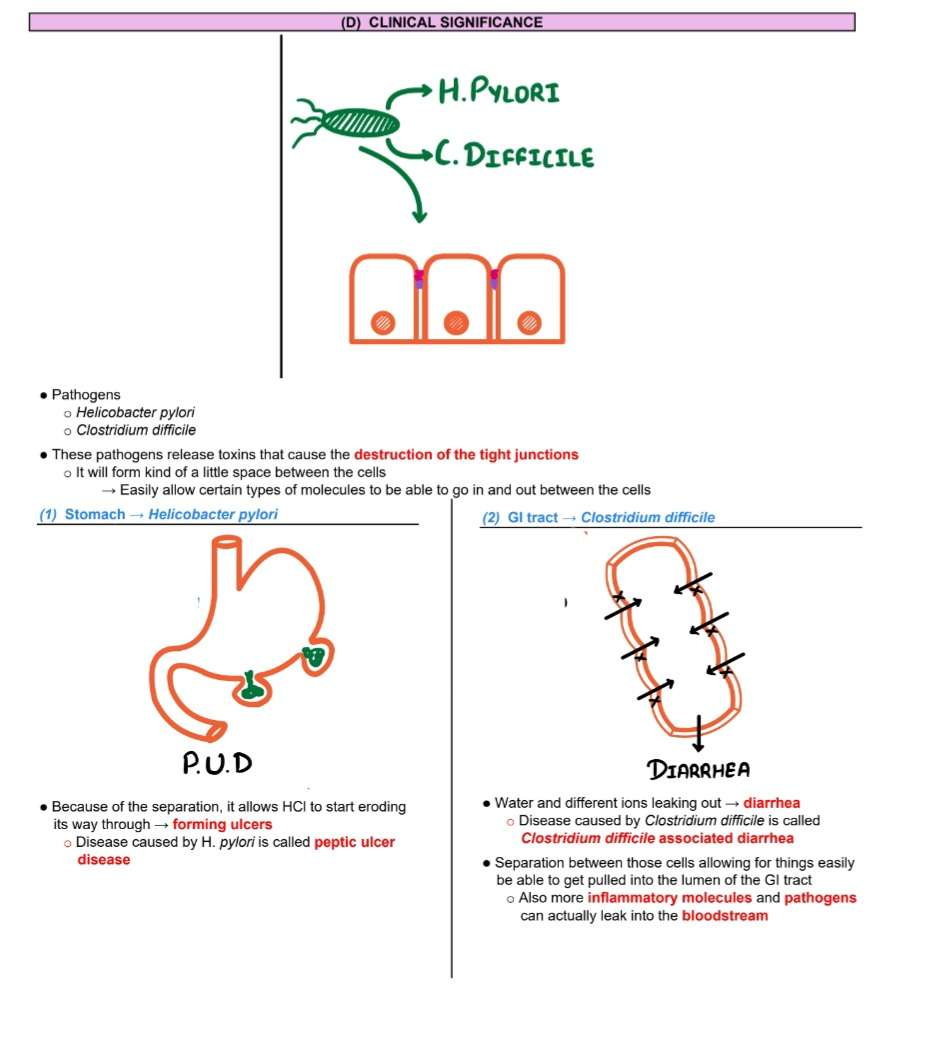
Test questions
Q1: The actual part of the protein that spans through the cell membrane out to the extracellular component and attaches cell to cell → A: Claudins and occludins
Q2: Proteins on the cytosolic side that is bound to the actual transmembrane proteins → A: Zona occludins
Q3: Proteins that are bound on the most inner cytosolic side to the zona occludins → A: Actin filaments
Adherence Junctions:
Definition: Cell-cell junctions that connect the actin cytoskeletons of neighboring cells.
Location: Found just below tight junctions in epithelial tissues.
Components:
Cadherins
Calcium-dependent proteins are critical for cell-cell adhesion and junction maintenance of tissue integrity.
Structure:
Key proteins: Cadherins (calcium-dependent adhesion molecules)
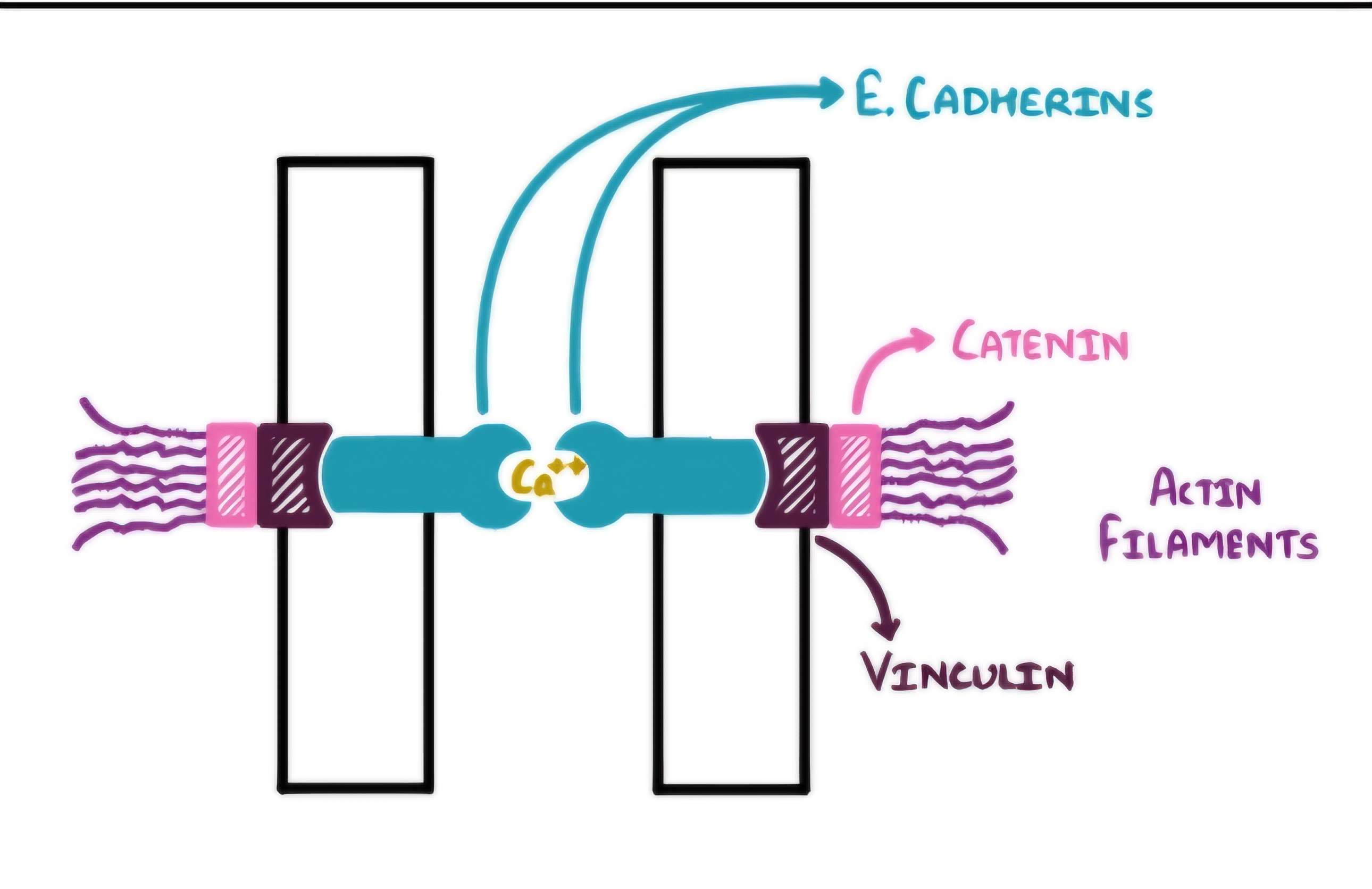
On the cytosolic sideLink to actin filaments via catenins and Vinculin
Cadherins link to actin through a trio of key proteins:
β-catenin – binds directly to cadherin tail
p120-catenin – stabilizes cadherin at the membrane
α-catenin – connects β-catenin to actin filaments and recruits actin-regulating proteins
Functions:
Binds to matching cadherins on adjacent cells (homophilic binding)
Anchors to actin filaments inside the cell via catenins (cytoplasmic linker proteins)
Transmits mechanical forces from cell to cell across tissues
Helps maintain tissue integrity during movements like stretching or morphogenesis
Exist in various forms, contributing to the stability of junctions by mediating strong binding between cells.
Maintains tissue integrity and mechanical strength.
Much stronger than a tight junction and are really good for shearing forces and abrasive forces
Made for a lot of abrasive forces.
Supports cell polarity, shape, and sorting during development
Important in morphogenesis and cell signaling.
Anywhere there are tight junctions, there are adherens junctions
Located:
just below tight junctions on the lateral sides of epithelial cells.
It anchors into the cell membrane, and the component then bulges out from the cell membrane and into the extracellular space.
Calcium acts as the bridge; it binds with other calcium.
A critical element for cadherin function facilitates the binding between cadherins and stabilizes these cell-cell junctions, ensuring the integrity and strength of tissue architecture.
Other Factors
Basal lamina: They are more basal than tight junctions.
Adherens Junctions & Contractile Forces:
Tension at the Junction:
Most adherens junctions are linked to actin filaments + non-muscle myosin II → forms a contractile bundle.
These bundles generate pulling forces on the junction.
This tension is not destructive — it’s essential for:
Assembling the junction
Maintaining junction integrity
Balance Is Everything:
Contractile force from one cell is matched by the other.
Prevents any one cell from yanking its neighbors — keeps tissue evenly spaced and stable.
Adherens Junctions Are Tension Sensors:
Junctions sense mechanical stress and respond dynamically.
If one cell gets more contractile, the junction enlarges, and the neighboring cell ramps up tension too.
They are dynamic tension sensors not just passive
Role of α-Catenin in Mechanotransduction:
Under tension, α-catenin changes shape:
Goes from folded → extended
Exposes a hidden binding site for vinculin
Vinculin joins the party → brings more actin to reinforce the junction
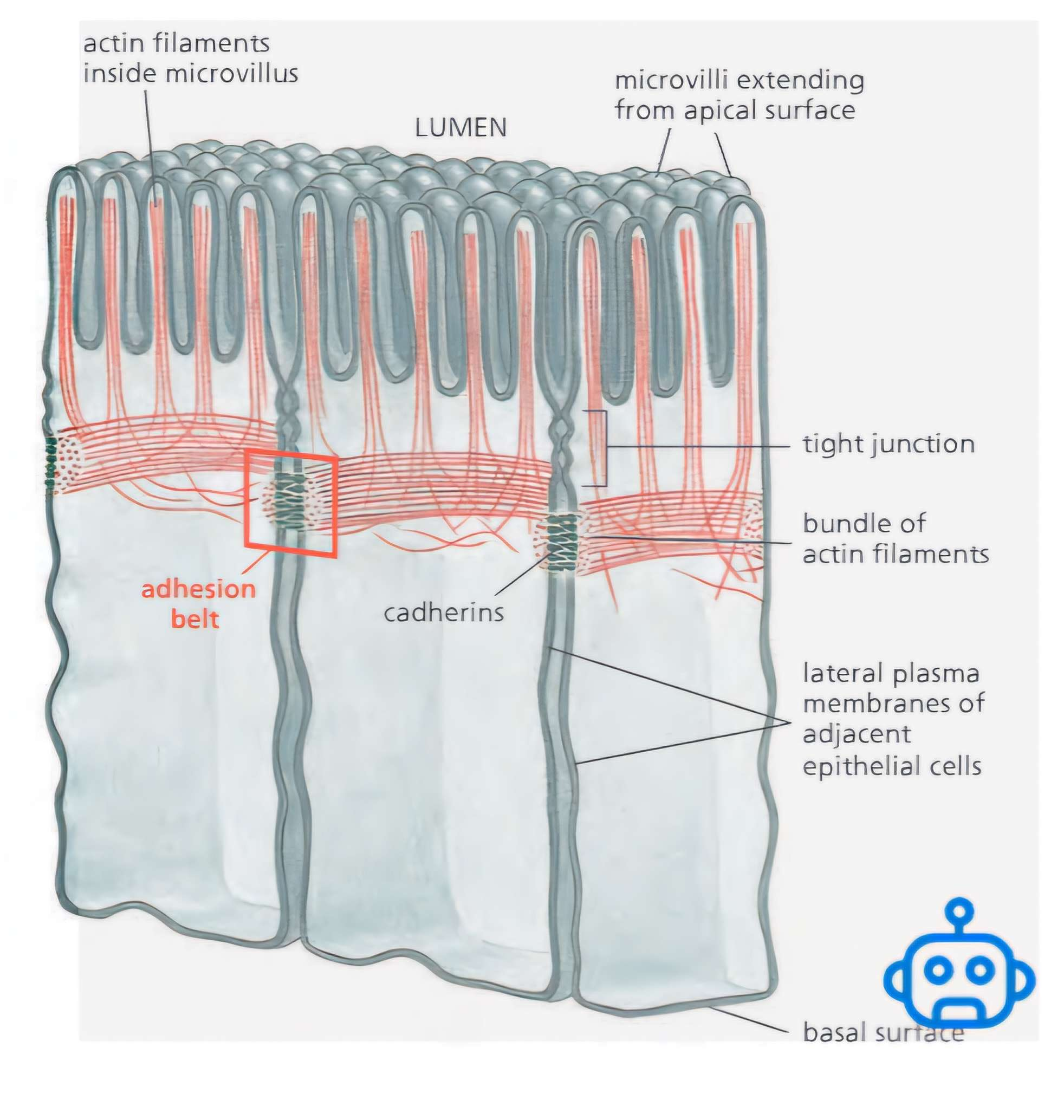
Adhesion belt
(or zonula adherens) just beneath the apical face of the epithelium, encircling each of the interacting cells in the sheet
Coordinated contraction of adhesion belts = motile force
Drives:
Folding of epithelial sheets into tubes
Formation of vesicles, organs, and other body structures
Dual Role of Force:
Small to moderate force = strengthens adhesion
Large force = can reduce adhesion, especially if:
Accompanied by regulatory signals that weaken cadherins
Seen in tissue remodeling and developmental processes
Too much pull, and the rope? 💥 Snaps (or detaches) — perfect for remodeling the field.
Clinical Ref
Heart muscle:Anchor contractile actin bundles; work with desmosomes to link heart cells end-to-end
GIT:
Respiratory tract:
Our lungs stretch and contract, so the use of adherence junctions is important for proper oxygen and carbon dioxide to be effectively exchanged.
Urinary tract Blood vessels:
The bladder needs to accommodate changes and stretches for urinary function
Skin: Stretches and abrasive forces
Cancer:
When cancer develops, mutated genes alter the structure of the Cadherin proteins. This results in the loss of their function, and the cells begin to separate in clumps. These clumps spread from a primary location to a secondary location in a process known as METASTASIS
Images



Desmosomes:
Definition: Spot-like cell-cell junctions providing strong adhesion.
Location: Prominent in tissues under mechanical stress (e.g., skin, heart).
Structure:
Proteins: Desmogleins, Desmocollins (cadherin family)
Spanning from the extracellular sides through the cell membrane into the extracellular matrix
They arch themselves like a zipper since there
Cadherins are between these structures and are calcium-dependent.
This intricate arrangement provides mechanical strength and stability to tissues, especially in areas subjected to stress, such as the skin and heart.
Structural cousin of adherens junctions
Appear as button-like “spot welds”
Intermediate filaments anchor to desmosomes on the intracellular side
These filaments form a continuous tensile network across cells
Anchored intermediate filaments (e.g., keratin) via plakoglobin and desmoplakin.
More inner cytosolic area, desmoplakin and plakoglobin interact with keratin to form a robust cytoskeletal framework, which is crucial in sustaining cellular integrity and facilitating communication between adjacent cells.
Keratin: Intermediate Filaments is the main component
Keratin filaments – in most epithelial cells
Desmin filaments – in cardiac muscle cells
Functions:
Provides mechanical strength.
Toughest and really high tensile against high shearing forces and abrasive stretch, these junctions are essential for maintaining tissue integrity and protecting cells from mechanical damage. Resists shearing forces.
How they work:
Desmogleins and desmocollins from adjacent cells bind to each other in the extracellular space (requires calcium).
Inside the cell, their cytoplasmic tails link to plakoglobin and desmoplakin, which anchor to intermediate filaments (like keratin).
This creates a strong, spot-weld-like connection that resists pulling and shearing forces.
Clinical Ref.
Essential in tissues exposed to high stress, like skin and cardiac tissue
Why:
Pemphigus Vulgaris: autoantibodies attack the desmoglein, and the cells of the epidermis separate. This usually involves the oral mucosa. It hinders desmosomes' ability to maintain structural integrity, leading to fragility and increased vulnerability to damage
Results in:
Severe blistering
ulcers
Leakage of body fluids
Potentially fatal if untreated
Images

Gap Junctions:
Definition: Communicating junctions allow direct transfer of ions and small molecules between adjacent cells. These are cell-cell junctions that facilitate intercellular communication, enabling synchronized activities such as muscle contractions and neuronal signaling.
How: that creates passageways linking the cytoplasms of adjacent cells via direct channels.
Channel Forming Junctions: Another name for Gap Junctions, it provides cytoplasmic connections through gap junctions, allowing for direct chemical communication between adjacent cells.
Not able: They are not diffusion barriers, resist abrasive forces, they don’t really connect cells to connective tissue
Location: Most tissues, especially cardiac and smooth muscle.
Channel Properties:
Pore size ≈ 1.4 nm.
Allows exchange of:
Inorganic ions
Small water-soluble molecules
Blocks larger macromolecules (e.g., proteins, nucleic acids).
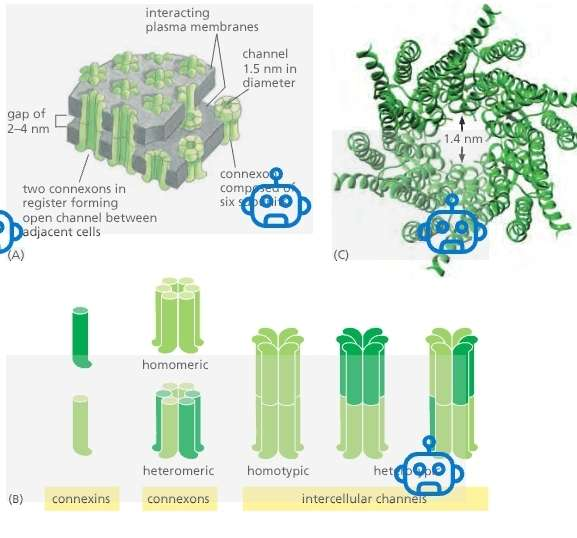
Structure:
Innexins:
found in invertebrates.
Connexons:
found in vertebrates (21 isoforms in humans).
Form the gap junctions
A gap junction has two connexons
Connexon + Connexon = Gap Junctions
Connexon pairing:
One connexon in each of two neighboring cells aligns perfectly.
Together, they form a continuous aqueous channel bridging the cytoplasm of both cells.
A gap junction consists of many connexon pairs in parallel, like a molecular sieve.
Connexins
Form the connexions
A connexon has six connexions
A gap junction has twelve connexions
Functions:
Enables cell-cell communication.
Important in electrical coupling and coordinated cellular activity.
Permeability
Regulation
Connexons can be:
Homomeric (same connexin type)
Heteromeric (mixed connexions)
Heterotypic channels: Each half-channel (connexon) is different across cells.
Key features:
Connexon plaques:
Can range from few to thousands of connexons.
Constantly remodeled.
Connexon turnover:
Half-life = a few hours (yep, they’re speedy recyclers).
New connexons are:
Added to the periphery of plaques via exocytosis.
Diffuse in the membrane and get trapped at the edge.
Old connexons are removed from the center
Unpaired Connexons:
Exist in membrane outside gap junctions.
Normally closed to prevent small molecule leakage.
In some cases, can open to release small signaling molecules.
Intracellular Ions:
Intracellular ions (e.g., sodium, calcium) can move from
one cell to another through gap junctions
Important in cells that are “excitable” (i.e., cells which
need to propagate electric potentials to nearby cells)
Not always open! They toggle between open and closed states, like ion channels.
Gated by:
Voltage differences between cells
Membrane potential
Cytoplasmic pH
Ca²⁺ concentration
Extracellular signals, e.g., neurotransmitters
Excitable cells include:
Electric current in one cell spreads to neighbor.
Has intercalated discs (composed of desmosomes
and gap junctions)
Smooth muscle tissue
Located along the gastrointestinal tract, respiratory
tract, urogenital tract, and blood vessels
Non-excitable tissues:
Enables sharing of small metabolites.
Helps coordinate activity and stabilize fluctuations in small molecule concentrations between cells.
Neurons
Some specific types of neurons communicate via
gap junctions
Key features
Allow electrical coupling (e.g., in cardiac and smooth muscle cells).
Enable chemical coordination (e.g., synchronizing cell responses).
Selectivity based on size and charge of molecules (~<1 kDa).
Regulated by pH, Ca²⁺, and voltage changes.
Cell Signaling
Cell signaling molecules (e.g., proteins, second
messenger molecules) to move from one cell to another
Cell signaling molecules include:
Cyclic AMP – activates protein kinase
A IP
When a ligand binds to the receptor on the cell
membrane, it can stimulate the cell signaling molecules
to travel through the gap junction to alert nearby cells, thereby facilitating rapid communication and coordination among neighboring cells.
Why this is important: Role in Pathology
When a cell is infected by a pathogen, it can alert the cell nearby to:
synthesize antimicrobial proteins against the pathogen
trigger apoptosis (programmed cell death)
trigger cellular adaptive processes (e.g., hypertrophy, atrophy)

Images

Hemidesmosomes:
Definition: Junctions that anchor epithelial cells to the basement membrane.
Location: Basal surface of epithelial cells.
Functions:
Provides stable adhesion to the extracellular matrix.
Maintains tissue architecture.
Structure/Compnents:
Cell-to-Matrix Junctions
These anchor cells to the extracellular matrix (ECM):
Hemidesmosomes
(Also) Focal adhesions – not mentioned earlier, but another key cell-to-matrix junction that connects actin filaments to ECM via integrins.
Basal Lamina (blue)
A layer of extracellular matrix is secreted by epithelial cells
Anchors epithelial cells to the underlying connective tissue
Made up of the following proteins: Fibronectin, Laminin, Collagen
Integrins (purple)
Integrins → found in hemidesmosomes and focal adhesions (cell-to-matrix), they are crucial for determining cell shape and motility, allowing cells to respond to their environment effectively.
Essential proteins that mediate attachment between cells and the ECM play a multifaceted role in cellular signaling and adhesion.
Spans the entire plasma membrane of cells and connects the cell to the extracellular matrix( basal lamina), facilitating communication and maintaining tissue integrity.
Capable of transmitting signals across cell membranes in both directions, allowing feedback mechanisms that inform cells about their surroundings
Outside the Cell:
Integrins (e.g., α6β4 integrin) bind to laminin in the basement membrane.
Inside the Cell:
The cytoplasmic tail of β4 connects to intermediate filaments (like keratin) via linker proteins such as plectin and BP230.
This stabilizes the structure—think of it like velcro between the cytoskeleton and the outside world.
Link to intermediate filaments (orange) inside the cell (mainly keratin).
Located along the inner cytosolic side of the cell membrane
Anchor the transmembrane proteins/integrins down
Include keratin
Basement Membrane
Hemidesmosomes form and maintain the connection
between the epithelial tissue and the basement
membrane.
This specialized structure provides essential support and anchorage, allowing for the proper organization of tissues and playing a crucial role in cell signaling.
Clinical Ref.
Dysfunctions of the hemidesmosomes affect epithelial tissues (e.g. skin, respiratory tract, GI tract, urogenital tract), most especially the skin
Attack: Certain diseases will attack the integrins that connect the basal lamina to the epithelial cells, which causes harm to the anchoring of the underlying extracellular matrix/ basement membranes, leading to compromised tissue integrity and function.

Image


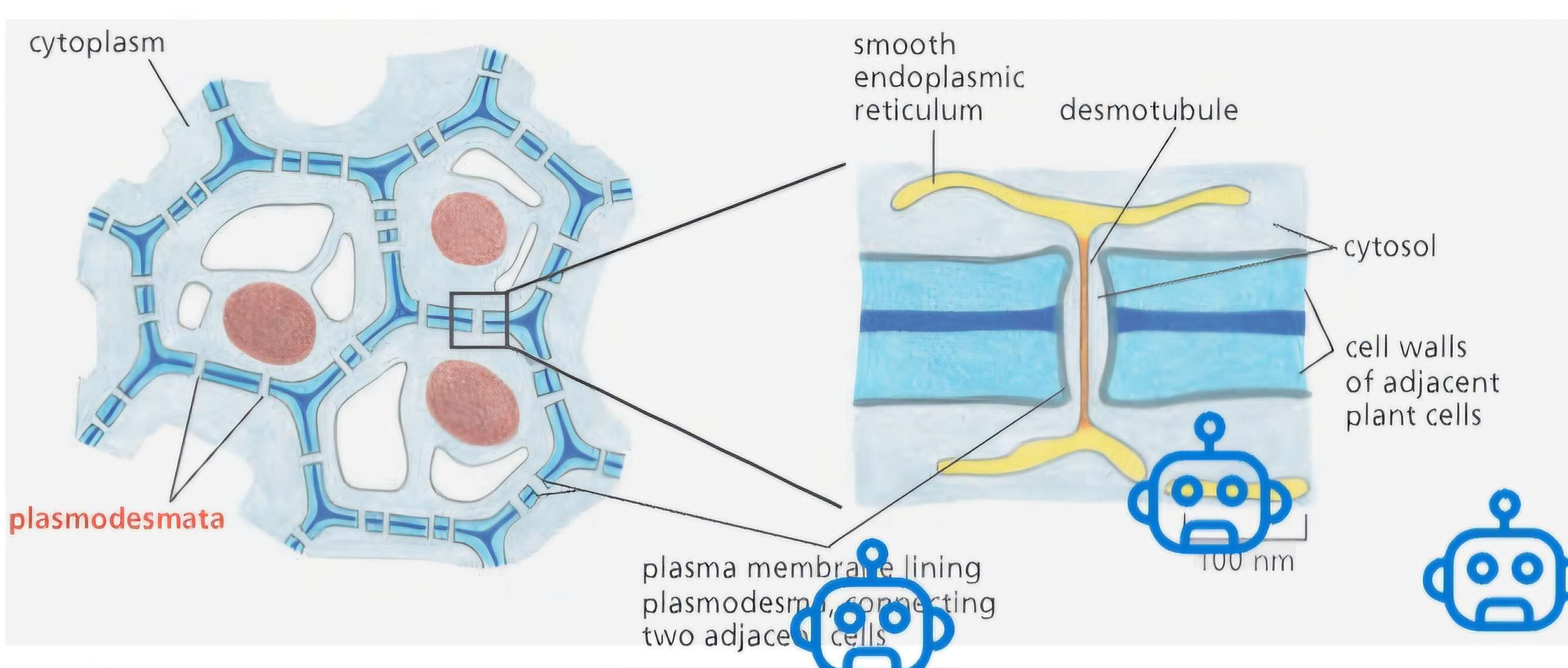
Plasmodesmata:
Purpose & Context
Plant cells are locked into place by tough cell walls rich in cellulose.
This strong wall removes the need for anchoring junctions.
But plants still need direct cell-to-cell communication.
Plasmodesmata = the only type of intercellular junction in plants.
They directly connect cytoplasms of adjacent cells—just like gap junctions!
Unique to plant cells, providing functionality similar to gap junctions but differing structurally.
Allow for cytoplasmic continuity and exchange of materials (e.g., small molecules, ions) through cell walls, facilitating communication between plant cells.
Structure of a Plasmodesma
Plasma membrane of neighboring cells is continuous at each plasmodesma.
Each plasmodesma is a cylindrical channel through the cell wall, ~20–40 nm wide.
Inside this channel:
A desmotubule runs through the center:
Narrow tube.
Continuous with the smooth ER of both cells.
Between the desmotubule and plasma membrane is a thin cytosolic sleeve:This is where small molecules pass from cell to cell.
Formation & Clustering
Plasmodesmata form:
During cytokinesis, when ER elements get trapped across the developing cell plate.
De novo, later on, inserted into mature cell walls.
Often found in clusters called pit fields.
Can be removed when no longer needed.
Function & Regulation
Functionally similar to gap junctions despite very different structure.
Molecular weight cutoff ≈ 800 Da, like gap junctions.
Regulated passage:
Some cells restrict movement through plasmodesmata, even to small molecules.
Mechanisms of restriction = not fully understood (mystery plants still keep 🌿).

Synapses:
Define neuron paths during development
Neurons use adhesion molecules to navigate to their correct destinations (a process called axon guidance).
Examples: N-CAM (Neural Cell Adhesion Molecule) and L1 guide growing axons through chemical and mechanical cues.
Think of them as GPS systems that lay down roads for neurons to follow.
Initiate synapse formation
When two neurons touch and their adhesion molecules interact, it triggers pre- and post-synaptic specialization.
This leads to the clustering of vesicle release machinery (presynaptic) and receptors (postsynaptic).
Molecules like neurexins (pre) and neuroligins (post) are matchmakers that handshake across the synaptic cleft.
Ensure proper alignment
Adhesion molecules ensure that neurotransmitter release sites line up perfectly with receptor clusters.
Without this alignment? Misfiring, weak signals, and possible neurological disorders (e.g., autism spectrum disorders have been linked to mutations in neurexins/neuroligins).
Hold neuron in place post-formation
After the synapse forms, adhesion molecules continue to maintain structural integrity.
They resist mechanical stress, keeping the neurons "locked in" for long-term communication.
Scaffold Proteins – The Synaptic Stage Crew
These are the master planners inside the synapse that make sure everything is exactly where it needs to be—no misplaced spotlight or mic stand here:
Organize junctional complexes
These complexes are like control centers at the synapse, full of machinery for signaling and plasticity.
Scaffold proteins (like PSD-95 in excitatory synapses) create a dense protein web that supports these zones.
Hold anchoring proteins
Anchoring proteins (like gephyrin in inhibitory synapses) attach receptors to the cytoskeleton or membrane.
Scaffold proteins hold and position these anchors exactly where they need to be.
Hold receptors and channels
Scaffold proteins bind to glutamate or GABA receptors, ion channels, and signaling molecules.
They ensure fast and precise signal transmission by clustering the right proteins at the right spot.
Think of them as event planners—everything’s scheduled and in place!
Supported by the cytoskeleton (actin)
The actin cytoskeleton provides a dynamic base, allowing the synapse to change shape during:
Plasticity (e.g., learning and memory)
Growth (e.g., during development)
Synaptic pruning or strengthening
Scaffold proteins link to actin, allowing the synapse to be both stable and adaptable.