Translation
Translation is the synthesis of a protein using the gene4tic information on the mRNA molecule.
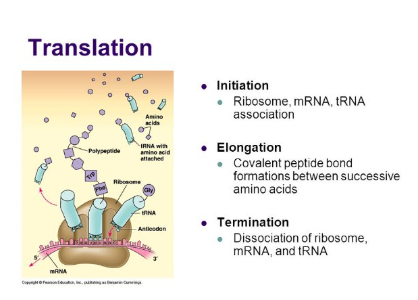
There are three steps in translation: Initiation, Elongation, and Termination
Translation takes place in the ribosome
Ribosomes are composed of two subunits: large and small subunit
Ribosomes are composed of proteins and rRNA
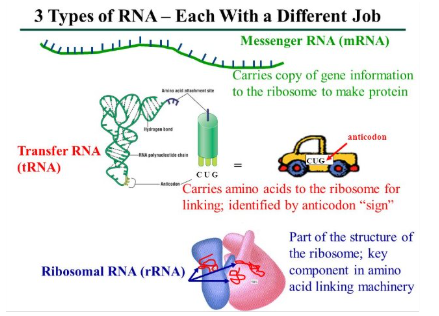
The three Types of RNA Involved in Translations:
Messenger RNA (mRNA)- carries information specifying amino acid sequences of proteins from DNA to ribosomes
Transfer RNA (tRNA)- interprets the information on a mRNA to add the proper amino acid to the growing protein chain
Ribosomal RNA (rRNA)- component of ribosomes
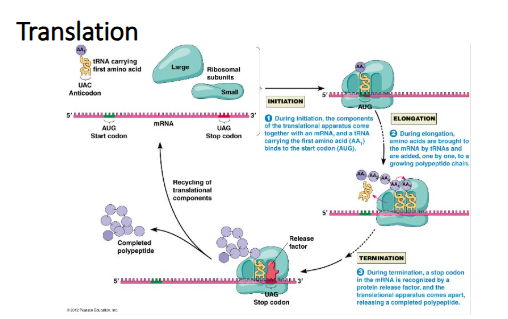
Step 1: Initiation
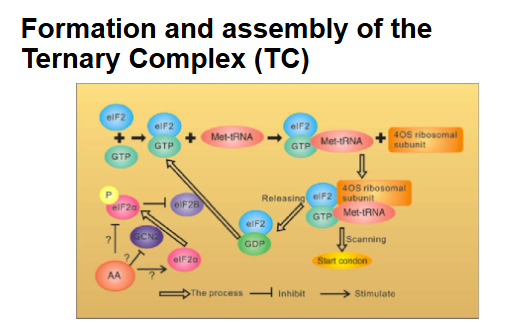
The first step of translation is the formation of the ternary complex (TC) which consists of: Initiator methionyl-tRNA (met-tRNAi), and the GTP bound form of the translation initiation factor elF2a
The TC then associates with the small (40s) subunit of the ribosome to form the 43S preinitiation complex
The eIF4E complex will bind to the 5’ 7mG cap and the elF4F complex consists of: elF4E which is the 5’ 7mG cap binding protein, elF4F complex which is an RNA helicase that unwinds the secondary structure of the 5’ UTR and elF4G which is a scaffold protein that links elF4E with the poly(a)- binding protein (PABP)
The 43S preinitiation complex will now bind to the elF4G on the 5’ UTR of the mRNA to form the 48S complex
The 40S ribosome with the bound translation initiation factors will scan the mRNA until it reaches the AUG start codon
Once the 40S ribosome reaches the AUG start codon, elF2a-GTP is converted into elF2a-GDP which will now result in all the bound translation initiation factors to be released of the 40S ribosome
The 60s (large ribosome subunit) will join with the 40S ribosome subunit to form the 80S initiation complex (IC) allowing for the elongation phase of translation to occur
The P site of the ribosome contains the initiator -tRNA bound to the first amino acid (Met) of the polypeptide chain
Step 2: Elongation
A new tRNA with an amino acid attached to it binds to the next codon on the mRNA and enters the A site of the ribosome
An enzyme on the ribosome call peptidyl transferase catalyzes a peptide bound between the two amino acids and the tRNA in the A-site will contain the dipeptide, which the initiator -tRNA in the P site now does not have an amino acid bound to it and now this initiator -tRNA will exit the ribosome through the E site
Now when the eukaryotic elongation factor 2 (eEF2) is converted from the GTP to GDP state ribosome moves or translocates to the next codon and now the tRNA with the growing polypeptide chain shifts from the A-site to the P-site and the next tRNA is ready to enter the A-site and this process continues until the ribosome reaches the stop codon
Step 3: Termination
When the ribosome reaches the stop codon, there is not a tRNA that will recognize the stop codon, therefore an animo acid is not deposited into the growing polypeptide chain
A translation factor known as eukaryotic release factor 1 (eRF1) will recognize the stop codon, which will result in the dissociation of the 80s ribosome from the mRNA and the polypeptide chain will be folded into a protein
Regulation of Translation:
Various environmental and intracellular stress such as: energy depletion, animo acid deprivation, hypoxia (low oxygen), misfolded proteins/aggregates, viral infections etc. can all lead to translation repression for a verity of ways
Two main pathways that regulate translation are: mTOR pathway and Regulation of ternary complex turnover (phosphorylation of elF2a)
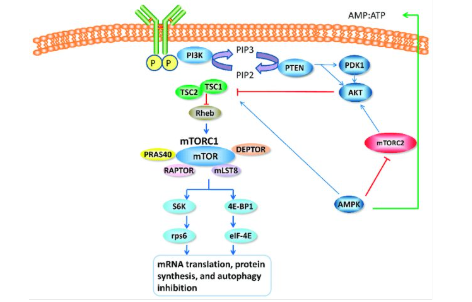
Regulation by mTOR:
Mammalian target of rapamycin (mTOR) is a kinase enzyme that regulates many pathways in eukaryotic organisms such as: cell proliferation, autophagy, and translation
During conditions in which nutrients are abundant, the mTOR pathway is activated, while depletion of nutrients will turn off the mTOR pathway
When the mTOR pathway is active it will promote translation in the following ways:
When mTOR is active it will phosphorylate the eIF4 inhibitory binding proteins (4E-BPs). The 4EBPs inhibit translation by sequestering eIF4E, therefore preventing eIF4E from being able to bind to the 5’cap. When 4EBP is phosphorylated, it cannot bind to eIF4E, therefore eIF4E is available to bind to the 5’ cap and allow for translation.
When mTOR is active it can phosphorylate ribosomal protein S6 (rpS6) allowing the rpS6 to associate with the ribosome to promote translation. If mTOR is inactive rpS6 is unphosphorylated meaning it will not associate with the ribosome, therefore slowing down translation.
When mTOR is active it will phosphorylate elongation factor 2 kinase (eEF2K). When eEF2K is phosphorylated, it will be inactive, and therefore eEF2K cannot phosphorylate eEF2. When eEF2 is unphosphorylated it is active and can promote elongation in translation allowing the ribosome to shift to the next codon.
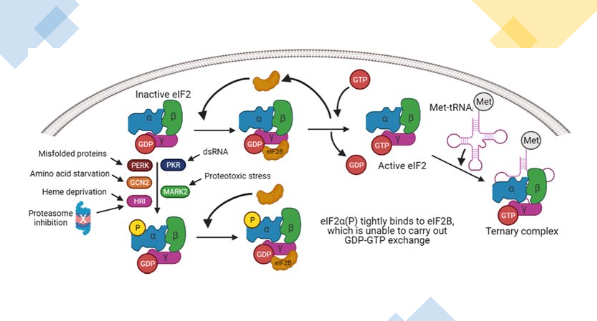
Regulation of ternary complex turnover (phosphorylation of elF2a)
Once the 40S ribosome reaches the AUG start codon, eIF2α-GTP is converted into eIF2α-GDP.
The regeneration of eIF2α-GTP is essential for the reassembly of the ternary complex (TC) so that continued translation can occur.
However, phosphorylation of eIF2α by various kinases: PERK ( activated by misfolded proteins, energy deprivation), GCN2 ( amino acid deprivation), PKR ( viral infections), HRI ( heme deficiency).
Phosphorylation of eIF2α causes tighter binding of eIF2B, which prevents eIF2B from exchanging the eIF2 α-GDP to eIF2 α-GTP, resulting in the inhibition of the initiation step of translation blocking the reassembly of the TC.