III. týždeň výv.
Tretí týždeň prenatálneho vývoja je kľúčový najmä vďaka procesu gastrulácie, počas ktorého sa pôvodne bilaminárne embryo premieňa na trilaminárne.
Vznikajú tak tri definitívne zárodočné listy: intraembryonálny ektoderm, entoderm a mezoderm
… pričom všetky tieto vrstvy sa diferencujú z pôvodného epiblastu.
Tieto zárodočné listy obsahujú multipotentné bunky, z ktorých sa následne vyvíjajú všetky tkanivá a orgány tela.
Gastrulácia zároveň umožňuje morfologické rozlíšenie hlavovej a chvostovej (kaudálnej) časti embrya, ako aj jeho pravej a ľavej strany.
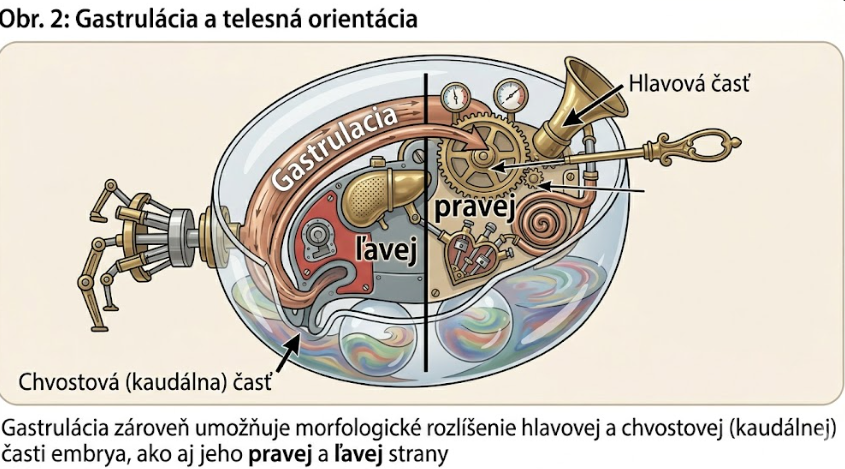
Z toho vyplýva ustanovenie bilaterálnej symetrie podľa strednej, respektíve hlavnej telovej osi.
Súbežne s gastruláciou prebieha notogenéza, teda proces vzniku troch osových štruktúr.
Časná notogenéza zahŕňa formovanie primitívneho prúžku a chordy dorsalis, zatiaľ čo neskorá notogenéza je spojená s vývojom neurálnej trubice.
Kľúčovou štruktúrou tohto obdobia je primitívny prúžok, ktorý vzniká proliferáciou buniek epiblastu v chvostovej časti embrya.
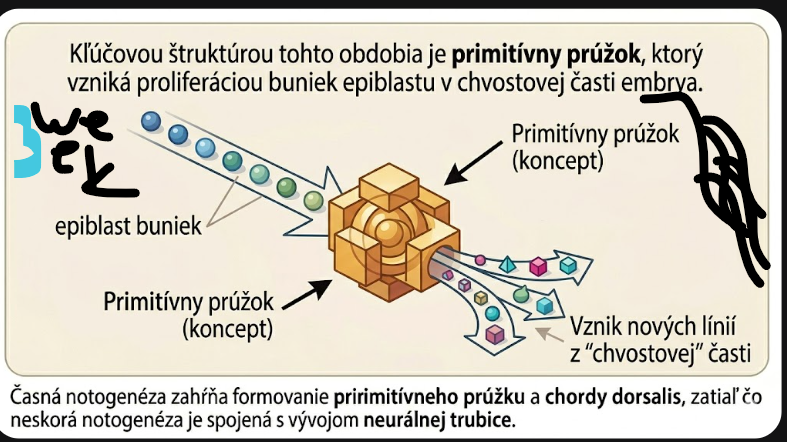
V jeho strede sa formuje proliferačná zóna označovaná ako primitívna brázda. Primitívny prúžok sa postupne predlžuje kaudo-kraniálnym smerom
a na svojom hlavovom konci sa rozširuje do primitívneho Hensenovho uzla, v ktorom sa nachádza primitívna jamka.
Prostredníctvom proliferácie a migrácie buniek z oblasti primitívneho prúžku sa tvoria nové zárodočné štruktúry. Prvá vlna migrujúcich buniek vcestováva do hypoblastu, roztláča ho laterálne a diferencuje sa na definitívny embryonálny entoderm. Zostávajúce bunky pôvodného hypoblastu, ktoré sú vytlačené na perifériu, následne tvoria extraembryonálny entoderm (ktorý tvorí strop žĺtkového vaku).
Po vytvorení entodermu nastupuje druhá migračná vlna, pri ktorej bunky prenikajú do priestoru medzi embryonálny ektoderm a novo sformovaný entoderm. Týmto procesom vzniká tretí zárodočný list – intraembryonálny mezoderm (mezoblast). Bunky epiblastu, ktoré nezačnú migrovať a zostanú na povrchu, napokon vytvoria (intra)embryonálny ektoderm. Je dôležité poznamenať, že tak embryonálny ektoderm, ako aj embryonálny entoderm si zachovávajú charakter epitelov a sú od vznikajúceho mezodermu obojstranne oddelené bazálnou laminou.
Vzniku samotného mezodermu (v III. týždni teda!!!) predchádza proces známy ako premena epitelových buniek na mezenchýmové (Epithelial-to-mesenchymal transformation, EMT).
Pôvodné epitelové bunky epiblastu, ktoré nasadajú na bazálnu laminu, strácajú v oblasti primitívnej brázdy svoj epitelový fenotyp. Prichádzajú o kontakt s bazálnou laminou, vycestovávajú zo steny primitívnej brázdy a menia sa na hviezdicovité mezenchýmové bunky. Časť buniek primitívneho prúžku si však zachováva svoj pôvodný fenotyp a naďalej plní proliferačnú funkciu. V prvej fáze produkuje primitívny prúžok bunky predovšetkým pre extraembryonálny mezoderm (mezenchým), ktorý sa podieľa najmä na stavbe zárodočného stvolu. Až keď je primitívny prúžok kompletne sformovaný, migrujú novovygenerované bunky priamo do štrbiny medzi epiblast a hypoblast, kde v špecifických lokalitách vytvárajú pruhy intraembryonálneho mezodermu. Tento sa anatomicky rozdeľuje na:
- paraaxiálny mezoderm (z ktorého vznikajú somity),
- intermediálny mezoderm (tvoriaci základ pre urogenitálny systém) a
- laterálny mezoderm, ktorý dáva vznik intraembryonálnemu coelomu. Z histologického hľadiska predstavuje mezenchým embryonálne pluripotentné tkanivo, z ktorého sa diferencujú všetky typy spojivových a podporných tkanív, hladká svalovina, krvinky a embryonálne väzivo.
Vývoj ďalších dôležitých štruktúr v osi embrya pokračuje formovaním chordového výbežku. Ten vzniká proliferáciou buniek z Hensenovho uzla, ktoré postupujú v strednej čiare medzi embryonálnym ektodermom a entodermom. Rast tohto výbežku smerom ku kraniálnemu pólu je priestorovo vymedzený a ukončený prechordovou platničkou. Táto platnička predstavuje okrsok vysokých cylindrických buniek hypoblastu na hlavovom konci embrya, ktoré tesne naliehajú na ektoderm. Prechordová platnička funguje ako dôležitý organizátor hlavovej časti embrya a zároveň vyznačuje oblasť budúcej orofaryngovej membrány. Na opačnom, kaudálnom konci embrya sa podobným mechanizmom formuje kloaková membrána. V mediálnej osi sa indukuje nielen chorda dorsalis, ale aj vznik neurálnej platničky (neuroektodermu), ktorá je priamym základom pre budúci nervový systém. Primitívny prúžok, ktorý všetky tieto transformačné zmeny inicioval a počas celého tretieho týždňa produkoval mezenchým, postupne stráca svoju funkciu a do konca štvrtého týždňa vývoja úplne zaniká.
Z epiblastu primitívneho prúžku proliferujú a migrujú bunky, ktoré formujú intraembryonálny mezoderm. Tieto bunky sa šíria rôznymi smermi a vytvárajú tak kaudálny, laterálny, intermediálny a paraaxiálny mezoderm. Najkraniálnejšiu časť intraembryonálneho mezodermu predstavuje mezoderm kardiogénnej zóny. Bunky tohto úseku migrujú z primitívneho prúžku kraniálnym smerom po oboch stranách strednej telovej osi, až do priestoru pred orofaryngovú membránu. Táto oblasť plynule nadväzuje na formujúci sa laterálny mezoderm, v ktorom sa následne tvorí dutina intraembryonálneho coelomu (ktorej súčasťou je v hlavovej časti aj perikardová dutina). Kardiogénna zóna poskytuje progenitorové bunky pre diferenciáciu kardiálnych myoblastov, z ktorých napokon vznikajú kardiomyocyty. Ďalšou dôležitou štruktúrou je zárodočný (úponový) stvol, tvorený extraembryonálnym mezodermom (mezenchýmom). V dôsledku rastu a ohýbania zárodku sa tento stvol presúva z vrcholu amniového vaku najprv do chvostovej časti a neskôr až na ventrálnu stranu embrya. Zárodočný stvol sa tak stáva priamou súčasťou pupočníka, pričom v ňom vznikajú umbilikálne cievy zabezpečujúce placentárny obeh.
V strednej osi embrya sa vyvíja chordový výbežok, ktorý sa formuje medzi 16. a 18. dňom vývoja. Vzniká proliferáciou mezodermových buniek z primitívneho Hensenovho uzla, ktoré vrástajú kraniálnym smerom medzi ektoderm a entoderm zárodočného terčíka, pričom výbežok rastie až k prechordovej platničke. Súčasne sa predlžuje aj primitívna jamka, čím vo vnútri výbežku vzniká chordový (Lieberkühnov) kanál. Spodná časť chordového výbežku následne zrastá s embryonálnym entodermom. Tieto spojené vrstvy sa vzápätí rozpadajú procesom apoptózy, čo vedie k vytvoreniu dočasnej komunikácie medzi amniovou dutinou a žĺtkovým vakom, označovanej ako canalis neurentericus. Tento prechodný stav umožňuje rýchlejšiu výmenu látok a živín prostredníctvom cirkulujúcej tkanivovej tekutiny. Rozpadom vzniká chordová platnička, ktorá sa prechodne laterálne spája s embryonálnym entodermom. Neskôr sa však od neho oddelí a vďaka bunkovej proliferácii sa ventrálne uzavrie, čím sa sformuje definitívna os embrya – chorda dorsalis. Táto štruktúra plní v časnom embryonálnom vývoji predovšetkým dôležitú indukčnú funkciu (napríklad pri vývoji tiel stavcov) a čoskoro degeneruje. Jej jediným pozostatkom, a to iba do časného detského veku, sú nuclei pulposi v medzistavcových platničkách.
Súbežne s vývojom osových štruktúr sa v laterálnom mezoderme a v mezoderme kardiogénnej zóny začínajú objavovať drobné dutinky. Tieto dutinky postupne splývajú a vytvárajú jednotnú embryonálnu telovú dutinu podkovovitého tvaru – intraembryonálny coelom. Spočiatku táto dutina široko komunikuje s extraembryonálnym coelomom, čo má zásadný význam pre výživu zárodku, prísun kyslíka a odvod katabolitov. Neskôr, v priebehu druhého mesiaca vývoja, sa táto pôvodne jednotná telová dutina rozdelí na tri definitívne priestory: nepárovú perikardovú dutinu vo vrcholovej časti, párové pleurálne dutiny vo forme kanálov a peritoneálnu dutinu, ktorá je spočiatku párová, no neskôr splynie do jednej.
Z hľadiska regionálnej diferenciácie sa intraembryonálny mezoderm po stranách chordy segmentuje na paraaxiálny mezoderm, z ktorého vznikajú somity (prvosegmenty, z ktorých sa neskôr diferencujú dermatom, myotom a sklerotom). Špecifickú nesegmentovanú kraniálnu časť paraaxiálneho mezodermu tvorí hlavový mezoderm. Ten sa mení na sieťovité tkanivo – hlavový mezenchým. Do tohto tkaniva neskôr vcestovávajú ektomezenchýmové bunky, vďaka čomu sa stáva zdrojom pre diferenciáciu kostrových svalov tváre, čeľustí a krku, čo úzko súvisí s vývojom faryngových oblúkov. Laterálne od somitov nadväzuje intermediálny mezoderm (základ napríklad pre mezonefros), ktorý plynule prechádza do platničky laterálneho mezodermu. Laterálny mezoderm sa čoskoro rozštiepi na dva listy: intraembryonálnu somatopleuru a intraembryonálnu splanchnopleuru. Práve tieto dva listy ohraničujú primitívnu jednotnú dutinu intraembryonálneho coelomu. Na laterálnych okrajoch zárodku tieto listy plynule prechádzajú do svojich extraembryonálnych proťajškov – extraembryonálna somatopleura sa nachádza na povrchu amnia a extraembryonálna splanchnopleura kryje žĺtkový vak. Týmto spôsobom je zabezpečená široká komunikácia dutiny intraembryonálneho coelomu s extraembryonálnym coelomom (choriovou dutinou).
Proces neurulácie, teda vývoj neurálnej trubice, sa začína približne v 18. dni vývoja a končí sa uzavretím zadného neuropóru (neuroporus posterior) v 28. dni. Východiskovou štruktúrou je neurálna platnička, ktorá predstavuje derivát embryonálneho ektodermu. Z neho sa vyčleňuje neuroektoderm, tkanivo s epitelovým charakterom. V neurálnej platničke sa formuje neurálna brázda (alebo ryha), po ktorej okrajoch sa dvíhajú neurálne valy. Tieto valy sú obzvlášť výrazné v kraniálnej oblasti, kde predstavujú priamy základ pre budúci mozog. Približovaním a spájaním neurálnych valov sa postupne formuje neurálna trubica a oddeľuje sa neurálna lišta.
Uzatváranie neurálnej trubice neprebieha synchrónne po celej jej dĺžke, ale začína od stredu embrya – v oblasti budúceho krku – a postupuje smerom k obom jej koncom. Predný otvor, označovaný ako neuroporus anterior, sa uzatvára medzi 25. a 26. dňom vývoja. Zadný otvor, neuroporus posterior, sa uzatvára o niečo neskôr, zhruba medzi 27. a 28. dňom. Z takto uzatvorenej neurálnej trubice sa následne vyvíja kompletný centrálny nervový systém, teda mozog a miecha.
Z neurálnej lišty, ktorá sa oddelila počas uzatvárania trubice, vzniká základ pre periférny nervový systém (PNS). Diferencujú sa z nej bunky spinálnych a vegetatívnych ganglií PNS, ako aj niektorých hlavových nervov (konkrétne V., VII., IX. a X. nervu). Okrem nervových buniek poskytuje neurálna lišta základ aj pre glie periférneho nervového systému, a to pre Schwannove a satelitné bunky. Neurálna lišta je však zdrojom aj pre mnohé nenervové deriváty. Vyvíjajú sa z nej všetky pigmentové bunky, dreň nadobličiek a ostatné paragangliá. Špecifickým derivátom je ektomezenchým. Vzniká tak, že multipotentné neuroepitelové bunky migrujú zo strednej partie neurálnej lišty a prostredníctvom epitelovo-mezenchýmovej transformácie (EMT) získavajú fenotyp mezenchýmových buniek. Tento ektomezenchým sa zásadne podieľa na formovaní mnohých štruktúr, vrátane chrupiek prvého až tretieho faryngového oblúka a ich derivátov v tvárovej a krčnej oblasti.
Od konca tretieho do začiatku piateho týždňa prebieha takzvaná somitová perioda, počas ktorej nastáva diferenciácia somitov (proces pokračuje až do konca 5. týždňa). Postupne sa v kraniokaudálnom smere vydiferencuje celkovo 42 až 44 párov somitov, z ktorých neskôr vznikajú ďalšie deriváty. Zaujímavosťou je, že na embryu nikdy nenájdeme všetky somity naraz. Vďaka ich presnej dynamike vývoja slúži v štvrtom a piatom týždni ich počet ako jedno z kritérií na určovanie veku embrya, známe ako Carnegieho štádiá. V treťom týždni sú diferencované prvé tri páry somitov a následne pribúdajú približne tri páry denne.
Samotnej diferenciácii definitívnych somitov predchádza vznik somitomér. Začiatkom tretieho týždňa sa najprv vydiferencuje 8 párov týchto štruktúr, ktoré sa vyznačujú jednoduchšou organizáciou ako somity. Somitoméry sa menia na mezenchým a podieľajú sa na formovaní niektorých kostí lebečnej klenby (kalvy) a bázy lebečnej. Z formovaných somitov neskôr vznikajú tri hlavné zložky. Zo sklerotomu sa vyvíja osový skelet, čo zahŕňa chrbticu, časť lebky, rebrá a hrudnú kosť (sternum). Dermatom dáva vznik zamši (dermis) a väzivu kože hlavy a trupu. Z myotomu sa formujú kostrové svaly vzťahujúce sa k osovému skeletu v oblasti hlavy, krku a trupu, vrátane epaxiálnej a hypaxiálnej svaloviny. Myotom predstavuje kľúčový zdroj buniek aj pre svalovinu končatín; myogénne prekurzorové bunky z tejto oblasti totiž priamo migrujú do vyvíjajúcich sa končatinových púčikov.