Chapter 11 Class Notes
Section 11.1 Monosaccharides Are the Simplest Carbohydrates
carbohydrates = carbon-based molecules high in hydroxyl groups
empirical formula: (CH2O)n
can have additional groups or modifications
better described as polyhydroxy aldehydes and ketones (and their derivatives)
Monosaccharides are aldehydes or ketones that contain two or more hydroxyl groups.
The smallest monosaccharides are composed of three carbons.
Monosaccharides exist in many isomeric forms.
Monosaccharides
monosaccharides = carbohydrates that are three to seven carbons in length
also called simple sugars
most common monosaccharides
4-7 in length
D or L sugars
based on glyceraldehyde
Monosaccharide Nomenclature
Nomenclature is based on carbon-chain length:
three carbons: trioses
four carbons: tetroses
five carbons: pentoses
six carbons: hexoses
fructose is a hexose
seven carbons: heptoses
Nomenclature is also based on the identity of the most oxidized group:
keto group: ketose
aldehyde group: aldose
Isomers
constitutional isomers
molecules with identical
molecular formulas that differ in how the atoms are ordered
different arrangement
stereoisomers
molecules that differ in spatial arrangement but not bonding order
have either D or L configuration
can be enantiomers (mirror images of each other) or diastereoisomers (not mirror images of each other)
number possible = 2n where n is the number of asymmetric carbon atoms
Fischer projections have the most oxidized group on top
Isomeric Forms of Carbohydrates
Common Monosaccharides
epimers = sugars that are diastereoisomers differing in configuration only at a single asymmetric center
Most Monosaccharides Exist as Interchanging Cyclic Forms
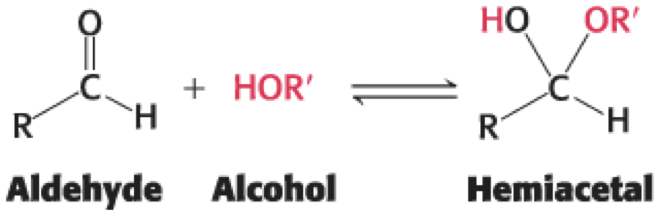
• An aldehyde can react with with an alcohol to form a hemiacetal
• A ketone can react with an alcohol to form a hemiketal
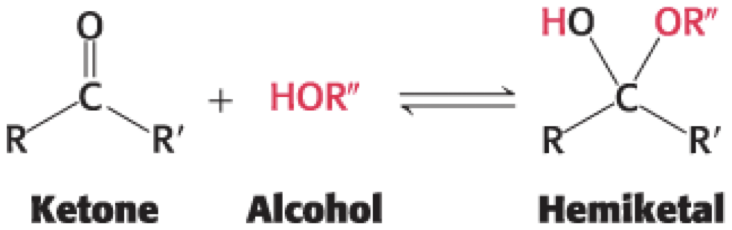
come from top: OH down
come from bottom: OH up
CH2OH and OH opposite → alpha
CH2OH and OH same → beta
fructose has 2 different CH2OH groups
Pyranose Formation
• called pyranose because of similarity to pyran
Furanose Formation
• called furanose because of similarity to furan
Anomers of Glucose
• anomer = a diastereoisomeric form of sugars that forms when a cyclic hemiacetal is formed and an additional asymmetric center is created (look at ring chains)
• In glucose, C-1 (the anomeric carbon atom) becomes an asymmetric center, forming two ring structures:
– α-D-glucopyranose (hydroxyl group attached to C-1 is on the opposite side of the ring as C-6)
– β-D-glucopyranose hydroxyl group attached to C-1 is on the same side of the ring as C-6)
D-Fructose Rapidly Interchanges Between Four Distinct Ring Structures
• C-2 is the anomeric carbon atom.
• The pyranose form predominates in solution due to reduced steric hindrances.
• The furanose form predominates in fructose derivatives.
The Most Common Monosaccharides Exist Primarily in Their Ring Forms
Pyranose and Furanose Rings Can Assume Different Conformations
• Pyranose rings are not planar because of the tetrahedral geometry of its saturated carbon atoms.
• They can adopt two types of conformation: boat and chair.
• In chair form, substituents on the carbon ring atoms can be axial (nearly perpendicular) or equatorial (nearly parallel).
• Axial substituents sterically hinder each other if on the same side of the ring.
Chair and Boat Forms of β-D-Glucose
• The chair form predominates because all axial positions are occupied by hydrogens.
• The boat form is disfavored because it is sterically hindered.
Envelope Conformations of Furanose Rings
• Furanose rings are not planar and commonly adopt a conformation called the envelope form.
• In the ribose moiety of most biomolecules, there are two common confirmations:
– C-2-endo (C-2 is out of the plane on the same side as C-5)
– C-3-endo (C-3 is out of the plane on the same side as C-5)
D-Glucose Is an Important Fuel for Most Organisms
• blood sugar = D-glucose circulating in the blood
– only fuel used by the brain in non-starvation conditions
– only fuel used by red blood cells
• potential reasons why D-glucose an important fuel:
– glucose is formed from formaldehyde under prebiotic conditions and may have been available as a fuel source for primitive biochemical systems
– glucose is relatively inert
– the most stable ring structure is β-D-glucopyranose
Solutions of Glucose
• The two anomeric forms (α and β) are in an equilibrium that passes through the open-chain form.
• There is a roughly 2:1 ratio of β-to-α anomer conformations for D-glucose in an equilibrium solution.
D-Glucose Is a Reducing Sugar and Reacts Nonenzymatically with Hemoglobin
• In its linear form, glucose can react with oxidizing agents.
• example: linear glucose reacts with Cu2+ yielding Cu+ and gluconic acid
Reducing Sugars
• Fehling's solution = solutions of Cu2+ that test for the presence of sugars that adopt an open structure
Cu2+ is easy to see
• reducing sugars = sugars that react with oxidizing agents (any sugar than can get to an open chain)
reducing agen is what is getting oxidized
– all monosaccharides that can adopt linear structures in solution
• non-reducing sugars = sugars that do not react with oxidizing agents
Glycation of Sugars
• glycation = nonenzymatic addition of a carbohydrate to another molecule
– can be benign or detrimental
• example: Reducing sugars nonspecifically react with free amino groups on proteins (often Lys or Arg) to form a stable covalent bond.
• D-glucose has a low tendency to glycate proteins unless concentrations of sugar and protein are very high for long periods of time.
Advanced Glycation End Products (AGEs)
• advanced glycation end products (AGEs) = products resulting from cross-linking following the primary modification
– implicated in aging, arteriosclerosis, diabetes, and other pathological conditions
Assessing Treatments for Diabetes Mellitus by Monitoring A1C Levels
• D-glucose reacts with hemoglobin to form glycated hemoglobin (hemoglobin A1c, A1C).
– has no effect on O2 binding
• In nondiabetic individuals, <6% of the hemoglobin is glycated.
• In patients with uncontrolled diabetes, almost 10% of the hemoglobin is glycated.
can be used as a makrer to look for diabetes
Monosaccharides Are Joined to Alcohols and Amines Through Glycosidic Linkages
• Monosaccharide biochemical properties can be modified by reactions with:
– alcohols.
– amines.
– phosphates → keep sugar in cell
• modifications increase biochemical versatility
– can serve as signal molecules
– can facilitate metabolism
sugars always modified
Glycosidic Linkages
• O-glycosidic linkage = covalent linkage formed between the anomeric carbon atom of a carbohydrate and the oxygen atom of an alcohol
• N-glycosidic linkage = covalent linkage formed between the anomeric carbon atom of a carbohydrate and the nitrogen atom of an amine
Monosaccharides Can Be Modified by the Addition of Substituents Other Than Hydroxyl Groups
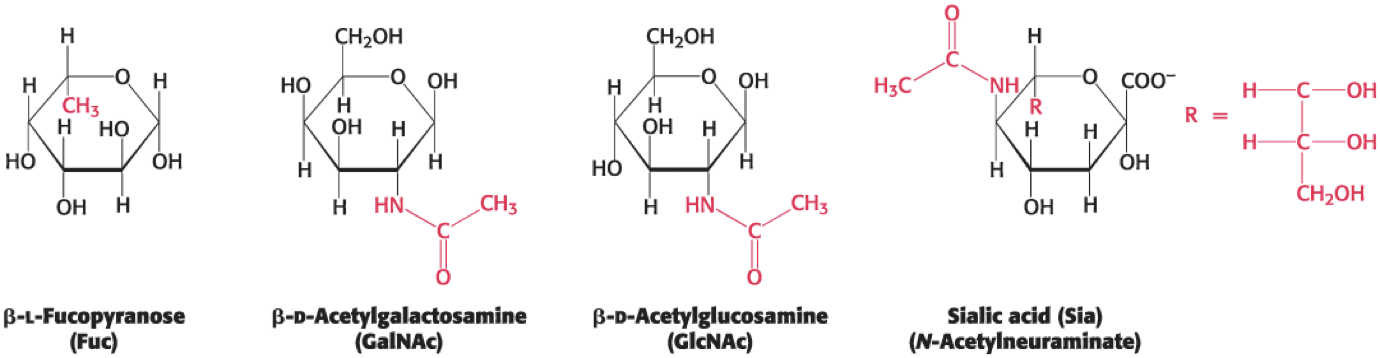 1 has added methyl
1 has added methyl
2 and 3 have added amid
used in siganl
not always at anomeric position
2 and 3 most important
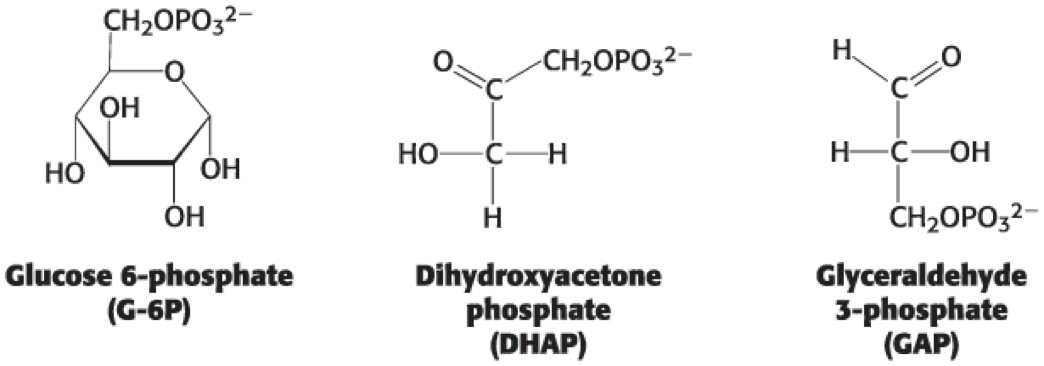
Phosphorylated Sugars Are Key Intermediates in Metabolism
• phosphorylation = addition of phosphoryl groups
– common modification of sugars in metabolic reactions
• purposes of phosphorylation:
– makes sugars anionic to prevent crossing the lipid-bilayer membranes and interacting with transporters of the unmodified sugar
– blocks the formation of alternative ring conformation
– creates reaction intermediates that more readily undergo metabolism
Several Intermediates in the Breakdown of Glucose Are Phosphorylated Sugars
Section 11.2 Monosaccharides Are Linked to Form Complex Carbohydrates
• oligosaccharides = sugars that contain two or more monosaccharides linked by O-glycosidic bonds
– have directionality defined by their reducing and nonreducing ends
• reducing end = has a free anomeric carbon atom that can form the open-chain form
• nonreducing end = has an anomeric carbon in a glycosidic linkage that cannot covert to the open-chain form
Maltose Is a Disaccharide of D-Glucose
• α-1,4-glycosidic linkage = glycosidic linkage between the α-anomeric form of C-1 on one sugar and the hydroxyl oxygen atom on C-4 of the adjacent sugar
Sucrose, Lactose, and Maltose Are the Common Disaccharides
• disaccharide = two sugars joined by an O-glycosidic linkage
• Cleavage products of disaccharides can be processed to provide energy in the form of ATP.
The Disaccharide Sucrose
• sucrose = disaccharide of sugar cane or sugar beets that consists of glucose linked to fructose
– the anomeric carbon of glucose is linked to the anomeric carbon of fructose
– the configuration is α for glucose and β for fructose
– not a reducing sugar
– can be cleaved by sucrase (invertase)
The Disaccharide Lactose
• lactose = disaccharide of milk that consists of a galactose linked to a glucose
– linked by a β-1,4-glycosidic linkage.
– can be hydrolyzed by lactase in human beings and by β-galactosidase in bacteria
The Disaccharide Maltose
• maltose = disaccharide resulting from the hydrolysis of large oligosaccharides that consists of two linked glucose molecules
– joined by an α-1,4-glycosidic linkage
– can be hydrolyzed to glucose by maltase (α-glucosidase)
Maltase Inhibitors Can Help to Maintain Blood Glucose Homeostasis
• After a meal, starch and glycogen are degraded by α-amylase.
• Oligosaccharides generated by α-amylase are further digested by maltase.
• Acarbose (Precose) and miglitol (Glyset) are competitive inhibitors of
maltase.
Glycogen and Starch Are Storage Forms of Glucose
• Free glucose cannot be stored because high concentrations will disturb the cell's osmotic balance.
• polysaccharides (glycans) = large polymeric oligosaccharides formed by the linkage of multiple monosaccharides
– plays roles in energy storage and structural integrity
• homopolymer = polymer in which all the monosaccharide units are the same
Glycogen
• glycogen = large, branched polymer of glucose residues
– most common homopolymer in animal cells
– storage form of glucose
– most glucose units are linked by α-1,4-glycosidic linkages
– branches are formed by α-1,6-glycosidic linkages
– hydrolyzed by α-amylase
• Branching increases the surface area to allow better access for enzymes to rapidly breakdown glycogen.
Starch
• starch = homopolymer that serves as the nutritional reservoir in plants
– two forms: amylose and amylopectin
• amylose = unbranched type of starch composed of glucose residues in α-1,4 linkage
• amylopectin = branched type of starch with ~1 α-1,6 linkage per 30 α-1,4 linkages
– identical structure to glycogen but with a lower degree of branching
• Amylose and amylopectin are hydrolyzed by α-amylase.
Cellulose Is the Main Structural Polysaccharide of Plants
• cellulose = unbranched polymer of glucose residues joined by β-1,4 linkages
– serves a structural role instead of a nutritional role
• The β configuration allows cellulose to form long, straight chains that interact with one other through hydrogen bonds
– yields a rigid, supportive structure
• The α linkages of starch and glycogen form compact hollow cylinders suitable for accessible storage.
Glycosidic Linkages Determine Polysaccharide StructureInsoluble and Soluble Fiber Are an Important Part of the Diet
• Mammals cannot digest cellulose because they lack cellulases, but plant fibers are still important in the mammalian diet.
• Insoluble fibers increase the rate at which digestion products pass through the large intestine.
– softens stools and makes them easier to pass
• Soluble fibers (e.g., pectin or polygalacturonic acid) slow the movement of food through the gastrointestinal tract.
– facilitates absorption of nutrients from the diet
Chitin Is the Main Structural Polysaccharide of Fungi and Arthropods
• chitin = homopolymer of β-1,4 linked N-acetylglucosamine
– found in fungal cell walls and exoskeletons and shells of arthropods
– Fibers are often crosslinked and composited with minerals and proteins to increase rigidity and strength.
Chitin Can Be Processed to a Molecule with a Variety of Uses
• Cellulose is a major constituent of paper, bioadhesives, and clothes.
• Chitin could be recovered from the shellfishing industry by processing the shells into the more versatile chitosan through microbial/enzymatic processes.
• Chitosan can be used as:
– a carrier to assist in drug delivery.
– a component of cosmetic and food products.
– a surgical dressing.
Section 11.3 Carbohydrates Can Be Linked to Proteins to Form Glycoproteins
• glycoprotein = a carbohydrate group covalently attached to a protein
– makes up 50% of the human proteome
• glycosylation increases the complexity of the proteome
– glycoforms = different glycosylated forms
– may occur when a protein has several potential glycosylation sites
Three Classes of Glycoproteins
• glycoproteins = predominantly proteins
– play a variety of roles, including cell adhesion
• proteoglycans = predominantly carbohydrates and the protein component is conjugated to a glycosaminoglycan
– function as structural components and lubricants
• mucins (mucoproteins) = predominantly carbohydrates and the protein components is extensively glycosylated at Ser or Thr residues, usually by N-acetylgalactosamine
– key component of mucus
– function as lubricants
Carbohydrates Can Be Linked to Proteins Through N-Linked or O-Linked
• N-linkage = links the sugars in glycoproteins to the amide nitrogen atom in the side chain of Asn
– Asn must be part of an Asn-X-Ser or Asn-X-Thr sequence, where X is any residue except proline
• O-linkage = links the sugars in glycoproteins to the oxygen atom in the side chain of Ser or Thr
N-Linked Oligosaccharides Have a Common Core
• N-linked polysaccharides have a common pentasaccharide core that consists of three mannoses and two N-acetylglucosamine residues.
The Glycoprotein Erythropoietin Is a Vital Hormone
• erythropoietin (EPO) = a glycoprotein secreted by the kidneys into the blood serum to stimulate production of red blood cells
– cloned recombinant form has improved treatment for anemia, but has been abused by some endurance athletes
– glycosylation enhances the stability of the protein in the blood
Oligosaccharides Attached to Erythropoietin
• N-glycosylated at three Asn residues
• O-glycosylated a Ser residue
• 40% carbohydrate by weight
Glycosylation Functions in Nutrient Sensing
• GlcNAcylation = the post-translational, covalent
attachment of a single N-acetylglucosamine (GlcNAc) to Ser or Thr residues of proteins
– catalyzed by O-GlcNAc transferase
– occurs when nutrients are abundant
– reversible
O-GlcNAc Transferase
• GlcNAcylation sites are also potential phosphorylation sites.
– O-GlcNAc transferase and protein kinases may be involved in cross talk.
• Improper regulation of O-GlcNAc transferase has been linked to:
– insulin resistance.
– diabetes.
– cancer.
– neurological pathologies.
Proteoglycans Have Important Structural Roles
• Proteoglycans are up to 95% glycosaminoglycan by weight)
– resembles a polysaccharide more than a protein
• Proteoglycans:
– function as lubricants and structural components in connective tissue.
– mediate adhesion of cells to extracellular matrix.
– bind factors that regular cell proliferation.
Glycosaminoglycans
• glycosaminoglycans = composed of repeating units of disaccharides containing a derivative of an amino sugar
– amino sugar derivative is either glucosamine or galactosamine
– at least one of the two sugars in the unit has a negatively charged carboxylate or sulfate group
• The inability to degrade glycosaminoglycans causes diseases marked by skeletal deformities and reduced life expectancies.
Glycosaminoglycans Are Made of Repeating Units
Proteoglycans Are Important Components of Cartilage
• Cartilage contains the protein collagen protein and the proteoglycan aggrecan.
• aggrecan = large molecule with three globular domains
– site of glycosaminoglycan (keratan sulfate and chondroitin sulfate) attachment is in the extended region between G2 and G3
– G1 noncovalently binds to a central polymer of hyaluronate
The Proteoglycan From Cartilage Has an Enormous and Complex Structure
Aggrecan Cushions Compressive Forces
• Water is bound to the glycosaminoglycans to cushion compressive forces.
– Water is squeezed from the glycosaminoglycan under pressure.
– Water rebinds when pressure is released.
• osteoarthritis = form of arthritis that results when water is lost from proteoglycan with aging
Mucins Are Glycoprotein Components of Mucus
• tandem repeats (VNTR) region = region of the protein backbone of mucins that is rich in O-glycosylated Ser and Thr residues
• Core carbohydrate structures are conjugated to the protein component of mucin.
Functions of Mucins
• Mucins:
– adhere to epithelial cells and act as a protective barrier.
– hydrate the underlying cells.
– play roles in fertilization, the immune response, and cell adhesion.
• Overexpression occurs in bronchitis, cystic fibrosis, and adenocarcinomas.
Protein Glycosylation Takes Place in the Lumen of the Endoplasmic Reticulum and in the Golgi Complex
• Endoplasmic reticulum (ER) and Golgi complex are organelles that play central roles in protein trafficking.
• N-linked glycosylation begins in the ER and continues in the Golgi complex.
• O-linked glycosylation occurs only in the Golgi complex.
Dolichol Phosphate
• dolichol phosphate = specialized lipid molecule located in the ER membrane
– contains about 20 isoprene (C5) units.
– location where large oligosaccharides destined for attachment to the Asp residues are assembled
– the terminal phosphate is the site of attachment
The Golgi Complex Is a Sorting Center
• Golgi complex = a stack of flattened membranous sacs
• proteins proceed to lysosomes, secretory granules, or the plasma membrane
– based on signals encoded within their amino acid sequences and three-dimensional structures
Specific Enzymes Are Responsible for Oligosaccharide Assembly
• glycosyltransferases = catalyze the formation of glycosidic linkages
• Activated sugar nucleotides are the most common carbohydrate donor for glycosyltransferases.
Blood Groups Are Based on Protein Glycosylation Patterns
• Blood groups are designated by the presence of one of the three different carbohydrates (A, B, or O) attached to glycoproteins and glycolipids on the surfaces of red blood cells.
• All blood groups have a core O antigen.
Specific Glycosyltransferases Add the Extra Monosaccharide to the O Antigen
• A and B antigens have one extra monosaccharide through an α-1,3 linkage to a galactose moiety of the O antigen
– added by specific glycosyltransferases
• type A transferase = adds N-acetylgalactosamin to form the A antigen
• type B transferase = adds galactose to form the B antigen
The A, B, and O Oligosaccharide Antigens Share a Common Core Structure
Blood Type Phenotypes Result from the Enzymes Present
• Individuals with the:
– O blood type lack both enzymes.
– AB blood type express both enzymes.
– A blood type express only type A transferase.
– B blood type express only type B transferase.
• have important implications for blood transfusions.
– If an antigen not normally present is introduced, the immune system recognizes it as foreign.
The Cholera Toxin
• cholera = disease caused by a toxin from Vibrio cholerae
• Individuals with blood type O are ~8 times more likely to have severe disease.
– The O antigen binds more tightly to the toxin than other blood type antigens.
Errors in Glycosylation Can Result in Pathological Conditions
• congenital disorders of glycosylation = pathological conditions resulting from improper modification of proteins by carbohydrates and their derivatives
– examples: certain types of muscular dystrophy are linked to improper glycosylation and I-cell disease
I-Cell Disease
• lysosomes = organelles that degrade and recycle damaged cellular components or endocytosed material
• I-cell disease = a lysosomal storage disease that causes severe psychomotor impairment and skeletal deformities
– affected lysosomes contain undigested glycosaminoglycans and glycolipids
– active enzymes responsible for degradation are synthesized
– enzymes lack appropriate glycosylation and are exported instead of being sequestered in lysosomes
• A mannose 6-phosphate residue of the N-oligosaccharide directs the enzymes from the Golgi complex to lysosomes.
• In I-cell disease, the mannose lacks a phosphate because patients are deficient in the N-acetylglucosamine phosphotransferase.
Biochemists Use Several Techniques to Analyze Oligosaccharide Components of Glycoproteins
• Oligosaccharides can be detached from the protein using enzymes that cleave oligosaccharides at specific linkages.
• Mass of an oligosaccharide can be determined using MALDI-TOF or other mass spectrometric techniques.
– Many possible oligosaccharide structures have the same mass.
Determining Oligosaccharide Structure and Points of Attachment
• Structure can be determined by combining additional cleavage of the oligosaccharide with mass spectrometry.
• Points of attachment can be determined by applying proteases to glycoproteins and performing chromatography.
– Fragments attached to oligosaccharides have chromatographic properties that with protease treatment.
– followed by mass spectrometry or direct peptide sequencing
• Oligosaccharides can be sequenced by using enzymes that cleave specific glycosidic bonds.
• MALDI-TOF is used to identify the released sugars.
Oligosaccharides Can Be Characterized by Mass Spectrometry
Section 11.4 Lectins Are Specific Carbohydrate-Binding Proteins
• glycan-binding proteins = bind to specific carbohydrate structures on neighboring cell surfaces
• lectins = class of glycan-binding proteins
– example: the mannose 6-phosphate receptor that binds and directs lysosomal enzymes to the lysosome
Lectins Promote Interactions Between Cells and Within Cells
• Lectins:
– function to facilitate cell–cell contact.
– usually contains 2+ carbohydrate-binding sites.
– are linked to carbohydrates by a number of weak noncovalent interactions.
Lectins are Organized into Two Large Classes
• C-type (calcium-requiring) lectins = found in animals
– function in receptor-mediated endocytosis and cell–cell recognition
• L-type lectins = rich in seeds of leguminous plants
– serve as potential toxins to herbivorous insects
– come act as chaperones in the eukaryotic ER
C-Type Lectins Use Calcium Ions to Bind Carbohydrates
• Ca2+ on lectin acts as a bridge between lectin and the sugar.
• Two Glu residues in lectin bind to Ca2+ and the sugar.
• Other hydrogen bonds form between lectin side chains and the carbohydrate.
Selectins
• selectins = members of C-type lectins
– bind immune-system cells to sites of injury in the inflammatory response
– play a role in recruiting leukocytes to inflammation sites
• different forms of selectins:
– L form = bind to carbohydrates on lymph-node vessels
– E form = bind to carbohydrates on endothelium
– P form = bind to carbohydrates on activated blood platelets
Influenza Virus Binds to Sialic Acid Residues
• hemagglutinin = influenza virus lectin protein that binds to carbohydrates sialic acid residues linked to galactose residues on cell-surface glycoproteins
– the virus is engulfed after binding
• The virus replicates inside the cell and viral particles bud off from the cell.
Neuraminidase Cleaves Oligosaccharide Chains
• Assembled viral particles are attached to sialic acid residues of the cell membrane by hemagglutinin.
• neuraminidase (sialidase) = influenza virus protein that cleaves the glycosidic linkages between sialic acid and the rest of the glycoprotein
– frees the virus to infect new cells
– inhibitors of neuraminidase (Tamiflu and Relenza) are
important anti-influenza agents
Influenza Virus Uses a Lectin for Specific Cell Binding