Structural Proteins
Characteristics of Structural Proteins:
Long, filamentous.
Generally insoluble.
Contains unusual amino acids.
Often contain cross-linked polypeptide chains.
Cellular:
Filamentous muscle protein.
Cytoskeletal proteins.
Keratin - epithelial cells.
Extracellular:
Collagen - connective tissue.
Elastin - yellow connective tissue (blood vessels).
Resilin - insect connective tissue.
Fibroin - silkworm silk.
Spidroin - spider silk.
Intracellular Structural Proteins:
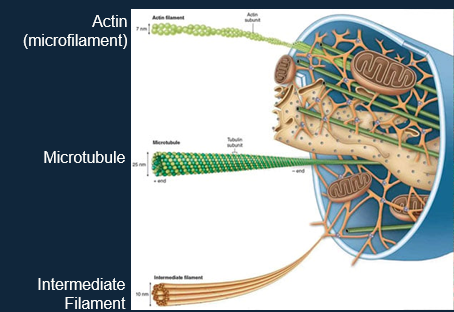
The cytoskeleton is composed of 3 types of structural protein:
— — — — — — — — — — —
Three Types of Cytoskeleton:
The overall function of the cytoskeleton is: endocytosis, cell division, intracellular transport, motility, reaction to external forces etc.
— — — — — — — — — — —
Actin Structure:
Two molecular types:
Globular (G-actin) - monomeric.
Filamentous (F-actin) - polymeric.
Filament formation:
G-actin + ATP - dimers or trimers (nucleation).
Elongation by monomer addition (treadmilling).
+ = barbed end.
- = pointed end.
Example of Actin Polymerisation and Regulation by ARP.
Binding of WCA (Wiskott–Aldrich syndrome protein (WASP)-homology-2, central, acidic) domain promotes a conformational change that primes the complex for activation, which occurs upon binding of the WCA–actin–ARP2/3 assembly to the mother filament, preferentially near the barbed end.
WCA domain presents an ATP–actin monomer to the complex and/or possibly to the barbed end of the mother filament.)
ATP is hydrolysed on ARP2 concomitant with or shortly after nucleation of the daughter filament.
The WCA dissociates, although the trigger for this is unknown.
Phosphate is released from ARP2. Mother and daughter filaments elongate and age by ATP hydrolysis and phosphate release.
Phosphate release from ARP2 and filament ageing weaken the interactions between ARP2/3 and the daughter and/or mother filament allowing branch disassembly and release of the ARP2/3 complex, presumably in an inactive, ADP-bound conformation.
Nucleotide exchange on ARP2 (and possibly on ARP3) occurs and the cycle begins again.
— — — — — — — — — — —
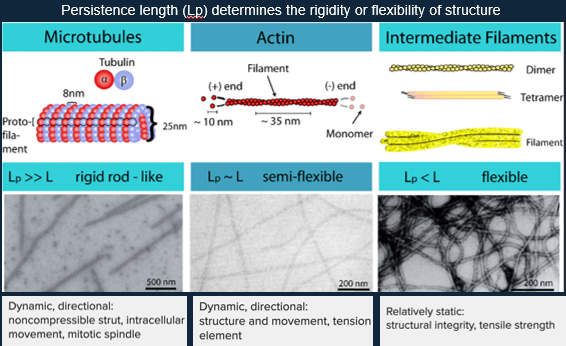
Microtubules:
Microtubules consist of α and β-tubulin monomers, and heterodimers.
Elongation:
Addition of heterodimers, forming a GTP-cap at the (+)-end, protecting microtubules from shrinkage.
Shrinkage:
If the (+)-end loses its GTP-cap, it induces microtubule shrinkage.
A single microtubule contains 10 to 15 protofilaments that wind together to form a 24nm wide hollow cylinder.
They are fueled by GTP hydrolysis.
Roles:
Major components of the cytoskeleton.
Found in all eukaryotic cells.
Involved in mitosis, cell motility, intracellular transport and maintenance of cell shape.
Microtubules tend to grow out from the centrosome to the plasma membrane.
— — — — — — — — — — —
Intermediate Filaments:
The intermediate filament (IF) supergene family are ubiquitous structural components that comprise the cell type-specific cytoskeleton of animal tissues.
All IF proteins show an organised, extended α-helical conformation prone to form two-stranded coiled coils.
Highly flexible, stress-resistant cytoskeletal filaments.
IF proteins are highly charged.
Keratin is the most common IF protein.
If has α-helices and β-pleated sheets.
α-keratins are intermediate filament proteins playing an important role in nuclei, cytoplasm, and cell surfaces.
E.g. in wool, they are mostly made of:
Glu/Gln - contribute to the hydrophilic properties.
Cys - essential for disulfide bond formation.
Ser - contains -OH groups making it hydrophilic.

α-keratin contains a coiled-coil (distorted right handed α-helix), a double α-helical coiled-coil with left handed supercoiling (dimer), a protofilament, protofibril, and an IF.
They contain cysteine bridges to stabilise the structure through covalent interactions.
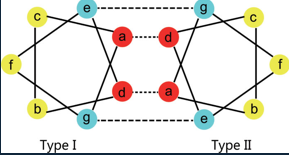
They have a heptad structure.
I.e. repeats of seven amino acids in positions labelled a-g. a-d are hydrophobic interactions, e-g are ionic H interactions.
a and d are often Leu, Ile, Ala.
e and g are often Glu and Gln.
Electrostatic interactions and salt bridges can form to enhance stability.
β-keratin is harder than its alpha counterpart.
Its’ monomer has hydrophobic interactions between 4β-strands, producing anti-parallel β-sheets.
Extracellular Matrix (ECM) Structural Proteins:
Collagen:
Makes up ~25-35% of all protein in the body.
Major component of connective tissues such as skin, tendons, cartilage, ligaments and bones.
Has a generalised amino acid composition:
33% Gly
11% Ala
10% Pro
5% Hydroxyproline (Hyp)
0.6% Hydroxylysine (Hyl)
0% Trp
Standard Structure of Collagen:

Molecular Structure of Collagen:
Glycine residues line the interior of the triple helix.
A lack of sidechains allows for tighter packing.
Bulkier side chains position on the outer faces of the helices.
Type I
Tendons, ligaments, bones.
Contains two α1 chains and one α2 chain.
Type II
Cartilage.
Contains three α1 chains (all three are identical).
— — — — — — — — — — —
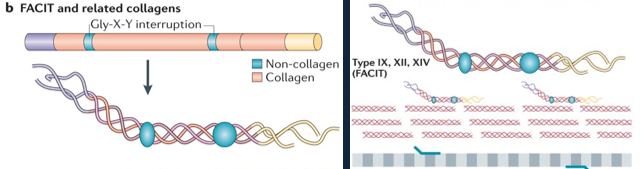
FACIT and Related Collagen Structures:
Fibril-associated collagens with interrupted triple helices (FACIT) and related collagens have a different structure to standard fibrillar collagen; they contain non-collagenous regions—that is, non-triple helical sequences.
These lead to kinks in the resulting macromolecular structure that straighten under small strains.
They associated with standard collagen, forming a higher-order structure.
— — — — — — — — — — —
Post-Translational Modifications:
Collagens are glycoproteins that are modified post-translationally.
Hydroxylation of Pro and Lys.
Glycosylation.
They are exported to the ECM (~1000 amino acids).
Deamination of Lys to aldehydes takes place, as well as cross-linking*.
— — — — — — — — — — —
*Cross-Linking of Collagen:
Collagens cross-link extensively post-translation (type I).
Lys is oxidised to ε-aldehyde (an Allysine intermediate), which is catalysed by LOX.
Condensation reactions take place, generating covalent cross-bridging between allysine:
Allysine;
Lysine (10)
Products: stable covalent bonds between LNL, DHLNL and HLNL.
Covalent cross-linking continues with time, which makes collagen more brittle with age.
— — — — — — — — — — —
Higher-Order Collagen Structures:
Triple stranded (3 separate chains) intertwined with a right-handed helical structure.
Staggered molecules.
Characteristic covalent cross-linking.
— — — — — — — — — — —
Collagen Defects:
Mutations in the collagen genes (~30) lead to severe disorders:
Osteogenesis imperfecta.
Osteoporosis.
Ehlers-Danlos syndrome.
Familial aortic aneurism.
Mutations at Gly-Xaa-Yaa lead to collagen misfolding, increased Lys hydroxylation, improper processing of protofilaments by enzymes and chaperones, and weakening of the collagen protofilaments.
General Structural Similarities between Silk and Spider Silk:
Relatively weak Van der Waals forces between sheets allowing for high flexibility.
The Gly-linked interface lacks large side chains and presents little resistance to bending.
Fibroin and spidroin also contain repeat polyalanine motifs.
These form crystalline domains which are held together by sericins.
This gives the fibre the strength of composites, and the flexibility of polymers.
— — — — — — — — — — —
Fibroin Compositions:
Fibrous protein secreted by insects and arachnids.
Amino acid composition of:
45% Gly, 30% Ala, 12% Ser, 5% Tyr.
Anti-parallel β-sheet.
— — — — — — — — — — —
Fibroin Structure:
Sheets, stabilised by hydrogen bonds, are stacked with side chains perpendicular to the structural plane.
Ala and Gly side chains are separated on opposite sides of the sheets.
Extended covalent structures allow little elongation and provide high tensive strength.
— — — — — — — — — — —
