Chapter 8: T Cell-Mediated Immunity
Activation of naïve T cells on encounter with antigen
T-cell activation or T-cell priming
The properties and functions of effector T cells
The interaction of effector T cells with their specific antigen that are presented by APC or target cells.
Naïve T cell action
First stage of a primary adaptive immune response
All effector T-cell differentiation happens in the periphery (secondary lymphoid organs) after antigen encounter.
Cytotoxic CD 8 T cells (kill infected cells)
CD4 T cells
Function = secrete cytokines activate other immune cells
Include:
TH1 - Treg
TH2 - TH17
TFH
Upon infection, the immune system quickly recruits the small number of naïve T cells that are specific to the pathogen make contact with antigens derived from the pathogen.
Recruitment involves secondary lymphoid tissues.
Antigen is brought in from outlying tissues via lymph.
Antigen encounters the T cells brought in via blood.
Examination of the activation of naïve T cells effector CD4 T cells by professional APC (P-APC) within lymphoid tissues
Dendritic cells
Interaction of a naïve T cell with antigen presented by cells other than P-APCs leads to inactivation rather than activation of T cells
Sentinel Dendritic Cells
The immune system does not initiate the adaptive immune response wherever a pathogen creates a site of infection.
The immune system captures the pathogen and takes it to the secondary lymphoid tissues.
Mediated by dendritic cells (migratory)
Process is same for infections in peripheral tissues, mucosal surfaces and blood
Dendritic cells are sentinels in all body tissues
Activated with pathogen uptake and antigen processing and presentation by MHC I and II molecules.
Dendritic cells – Major function is the triggering T-cell responses highly specialized and effective.
Dendritic cells are migratory cells and have highest concentration of B7 carry load of antigen from infection site nearest secondary lymphoid tissues.
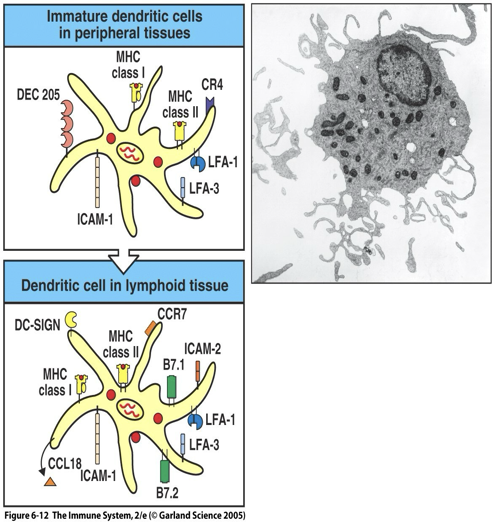
Movement of dendritic cell from site of infection in the periphery to a secondary lymphoid organ causes changes in the dendritic cell’s surface molecules, functions and morphology.
In tissues, dendritic cells are active in the capture, uptake and processing of antigen – called immature dendritic cells.
In secondary lymphoid tissues, dendritic cells gain the capacity to interact with T cells – called mature or activated dendritic cells (no longer phagocytic).
Mature dendritic cells have finger-like processes called dendrites that contact T cells in the cortex of lymph node.
Presenting to naive T cells
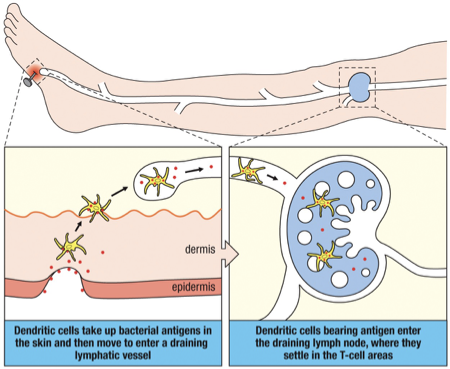
Wound in the skin.
Langerhans’ cells (immature dendritic cells) take up the antigen and migrate to a nearby lymph node.
Settle in the T-cell areas.
Differentiate into mature dendritic cells.
Immune Response Start Location
Skin and peripheral tissue infection = T cell response in draining lymph nodes.
Blood infections = antigen enter the spleen.
Respiratory mucosa infection = tonsils or other bronchial-associated lymphoid tissue (BALT).
Gastrointestinal infections = Peyers patches, appendix or gut-associated lymphoid tissues (GALT).
Similar sequence of events in each case.
Dendritic Cells Processing and Presentation
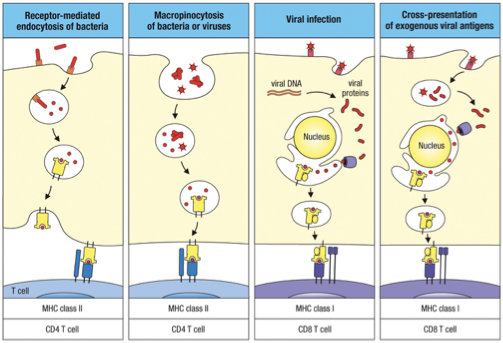
Immature dendritic cells use receptor mediated endocytosis to bring in microbial antigens using DEC 205 receptors (among others).
Antigens can also be taken up nonspecifically by macropinocytosis in which a cell engulfs extracellular fluid (ECF).
Dendritic cells acquire viral antigens via infection or by taking up virus particles from the ECF or other infected cells.
MHC I/CD8 T cell:
Viral infection antigen from inside
Cross-presentation of exogenous viral antigen
MHC II/CD4 T cell:
Receptor-mediated endocytosis
Macropinocytosis of bacteria or viruses
Changing function of Dendritic Cells
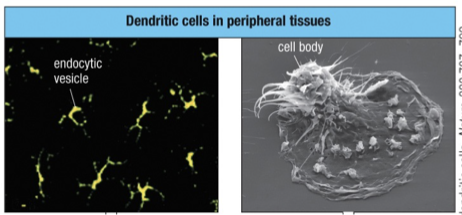
MHC class II stained green and a lysosomal protein (antigen) is stained red.
Cell bodies are difficult to discern
Dendrites contain endocytic vesicles that stain both for MHC class II & lysosomal protein giving rise to yellow fluorescence (combination of red and green stain).
On activation and migration in the lymph to secondary lymphoid organs the morphology of the dendritic cell changes.
Dendritic cells stop phagocytosis indicated by a partial separation of MHC class II (green) from the lysosomal protein (red)
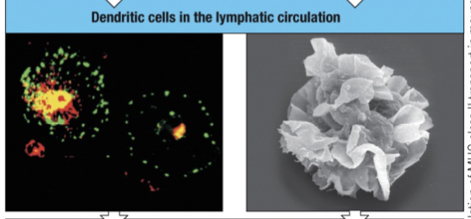
On reaching a lymph node “mature” dendritic cells begins antigen presentation to T-cells, stimulating the T cells instead of uptaking and processing antigen.
Lysosomal protein (red) is distinct from the MHC class II molecules (green).
Settles in T cell area, surface loaded with MHC class II and B7
MHC class II (green) is displayed at high density on the many dendritic processes.
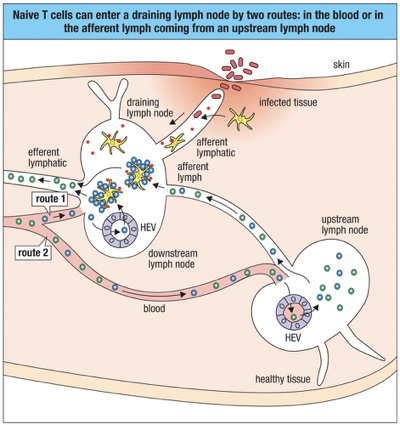
Circulation of Naive T cells
Naïve T cells enter lymphoid tissues via blood capillaries or from the afferent lymph.
Naive T Cells Entering Lymphatics:
Naive T cells enter lymph nodes through high endothelial venules (HEVs).
Chemokines released in HEVs: CCL19 and CCL21 attract naive T cells with CCR7 to lymph nodes.
T cells then interact with mature dendritic cells
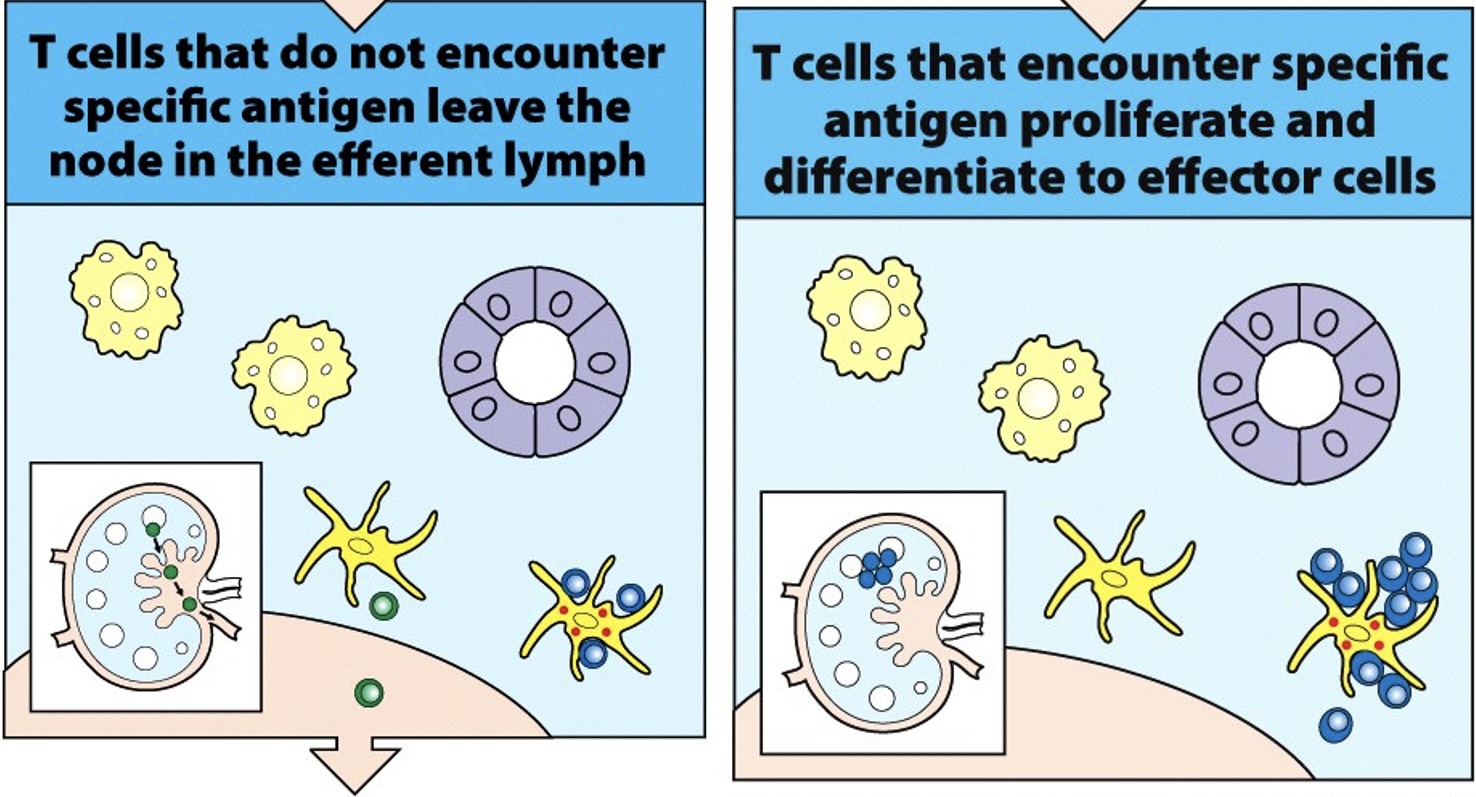
T cells that do not encounter their specific cells and antigen (green) leave the lymph node in the efferent lymph and eventually rejoin the bloodstream.
T cells that encounter antigen (blue) on antigen-presenting cells are activated to proliferate and to differentiate into effector cells.
These effector T cells can also leave the lymph node in the efferent lymph and enter the circulation.
Once an antigen-specific T cell is trapped in the lymph node by an Antigen Presenting Cell or APC (i.e Dendritic Cell) and activated it takes several days for the activated T cell to proliferate and differentiate into effector T cells .
delay between onset of infection and appearance of primary adaptive immune response.
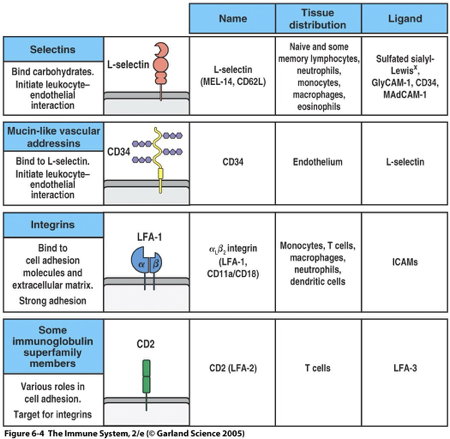
Leukocyte Adhesion Molecules
The four structural classes of adhesion molecule present on white blood cells and the cells with which they interact are:
Selectins – are carbohydrate-binding lectins; L-Selectin
Vascular addressins – contain carbohydrate groups to which selectins bind; CD34, GlyCAM, MadCAM
Integrins; LFA-1
Proteins in the immunoglobulin superfamily; ICAM (1,2,3), CD2, LFA3, CD28, B7
Binding rules:
selectins to addressins
Integrins to immunoglobulin superfamily (I to I)
binding can occur within a group as well
Homing
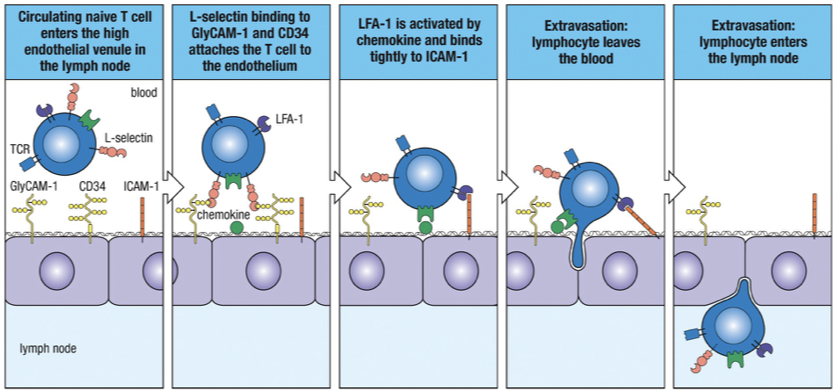
Movement of naïve T cells into secondary lymphoid tissues determined by T-cell surface selectin (L-selectin) interacting with two vascular addressins on surface of HEV venules (CD34 & GlyCAM-1)
L-selectin CD34 + GlyCAM-1
Tight Binding
Chemokines, which are also bound to the endothelium, activate the integrin LFA-1 on the lymphocyte surface enabling it to bind tightly to ICAM-1 on the endothelial cell.
LFA-1 ICAM-1
Diapedesis
Establishment of tight binding allows the lymphocyte to squeeze between two endothelial cells, leaving the lumen of the blood vessel and entering the lymph node.
The T-cells will migrate toward the high concentrations of CCL21 and CCL19 within the lymph node.
Immunoglobulin superfamily molecules adhesion to APC (Dendritic Cell)
As naïve T cells move through the cortex of the lymph node, these will bind transiently to APCs.
The initial encounter of T cells with antigen-presenting dendritic cells involves integrins and members of the immunoglobulin superfamily:
The T cell’s LFA-1 binds to ICAM-1 or ICAM-2 on the APC
The APC’s LFA-1 binds to ICAM-3 on the T cell
Adhesion is strengthened by intrafamiliar interactions of the immunoglobulin superfamily molecules:
CD2 on the T cell binding to LFA-3 on the APC.
ICAM-3 on the T cell and the lectin DC-SIGN on activated dendritic cell.
Costimulation with CD28 on T Cell and B7 on APC
These transitory interactions allow the T cell to screen the peptide:MHC complexes on the APCs (signal 1)
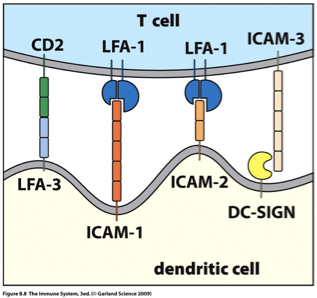
Memory trick;
LFA1 on T cell surface and always binds to ICAM1/ICAM2 (must be on dendritic cell surface)
LFA-3 on opposite side of LFA-1 (on dendritic cell surface) and binds to CD2 (must be on the T cell surface)
DC-Sign; DC for dendritic cell surface and always binds to ICAM-3 (must be on T cell surface)
Antagonist Adhesive Interactions
When a naïve T cell binds to its specific ligand on an antigen-presenting dendritic cell conformational change in LFA-1 that causes it to bind with higher affinity to ICAMs on the antigen-presenting cell suppress expression of S1P receptors T cell proliferates and differentiates into effector cells.
If antigen isn’t met, then the T-cells will be drawn out of the lymph node by a different chemotactic molecule called sphingosine 1-phosphate (S1P).
Now the T-cell can continue to recircluate until it meets its antigen.
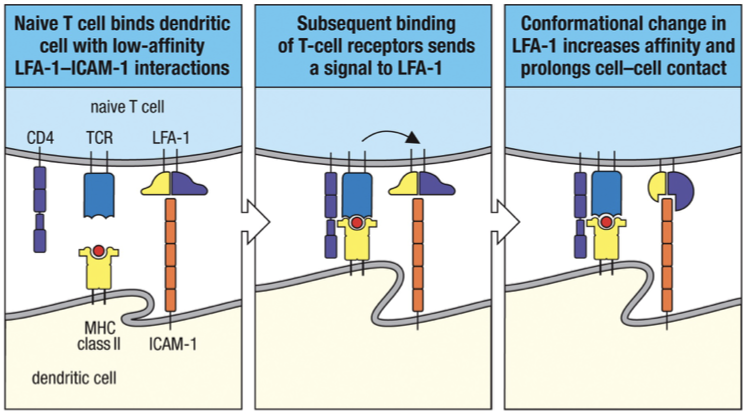
T Cell Signal Activation
Activation of naïve T cells CD28 binding to B7 along with peptide:MHC complexes binding to TCR and co-receptor binding.
Signal 1:
Binding of T cell receptor (TCR) and co-receptor CD4 to specific peptide-MHC complex on antigen-presenting cells (APCs).
TCR binds to MHC II and peptide (antigen)
CD4 co-receptor binds to non-polymorphic (constant) region of MHC II
Signal 2 (Co-stimulation):
Necessary for full T cell activation; involves interaction of B7 molecules on the APCs/dendritic cell with CD28 on T cells.
Without this signal, T cells may become anergic or undergo apoptosis.
B7 expression is a direct consequence of infection, induced by interaction of a potential APC with microbial products via innate receptors (TLR’s, mannose receptors…) that recognize pathogenic molecules (LPS, ds DNA, viral RNA…)
Therefore in absence of infection professional APC does not express co-stimulatory molecules
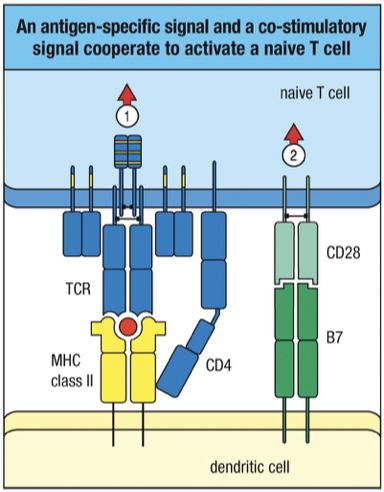
Process (homing, adhesion, and 2-signal activation) is the same for CD4⁺ and CD8⁺ T cells; Difference is that CD4 binds MHC class II while CD8 binds MHC class I, each using its co-receptor to attach to the non-variable part of the MHC.
Activated T cells express another B7 receptor called CTLA-4.
CTLA-4 binds B7 twentyfold more strongly than does CD28 and functions as an antagonist.
B7 binding to CD28 activates a T cell
B7 binding to CTLA-4 slows down activation and limits cell proliferation.
APC Characteristics
The characteristic that distinguishes professional antigen-presenting cells (APCs) from other antigen-presenting cells is the presence of B7 co-stimulatory molecules on their surface.
The three kind of professional APCs are: the dendritic cell, the macrophage and the B cell.
The one cell we know can activate naïve T-cells is the Dendritic cell.
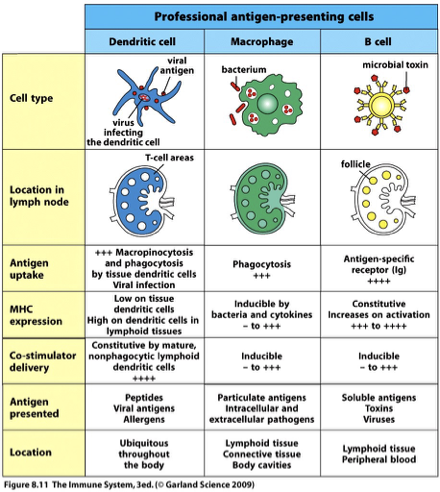
Locations;
B cell in follicle
T cell in T-cel area
Macrophages all throughout
Antigen uptake;
B cell through antigen
Dendritic Cells
Immature versus Mature Dendritic Cells (DCs):
Immature DCs express lower levels of co-stimulatory molecules and are more effective in capturing antigens.
Mature DCs express high levels of MHC and co-stimulatory molecules, promoting T cell activation.
On skin infection, local Langerhans’ cells will take up and process microbial antigen before traveling to the T-cell cortex areas of the draining lymph node and maturing to become a professional APC.
Activated (mature) dendritic cells not only express B7 and class II MHC molecules but also express high levels of adhesion molecules (i.e. DC-SIGN) and secrete the chemokine CCL18 specifically attracts naïve T-cells in peripheral tissue.
DC-SIGN binds to ICAM-3 on the T-cell
Immature DC:
DEC205 facilitates receptor mediated endocytosis and pinocytosis of antigens.
-ex: langerhans cells
Mature DC:
DC-SIGN uncrease expression of B7 co-stimulators, MHC molecules and adhesions molecules
-ex: interdigitating reticular cells
Macrophages
Macrophages are found throughout the lymph node tissue and have several different functions:
Are phagocytic cells that take up microbes and particles from the extracellular environment.
Degrade microorganism in phagolysosomes that are loaded with hydrolytic enzymes.
Trap and degrade pathogens this enables macrophages to process and present antigen to prevent infection from reaching blood.
Prevents noninfectious particulates from lymph nodes from entering blood and blocking small blood vessels.
Remove and degrade lymphocytes that die in secondary lymphoid tissues.
B-Cells
B cell Ig binds specific antigen from extracellular environment.
Ag:Ig complex internalized by receptor-mediated endocytosis transported to endocytic vesicles degraded into peptides peptides bind MHC class II molecules peptide:MHC class II complex transported to cell surface
In the primary immune response, the naïve B-cells are activated by effector T-cells that were activated in the same secondary lymphoid tissue.
Clustering of TCR and Signal Pathways
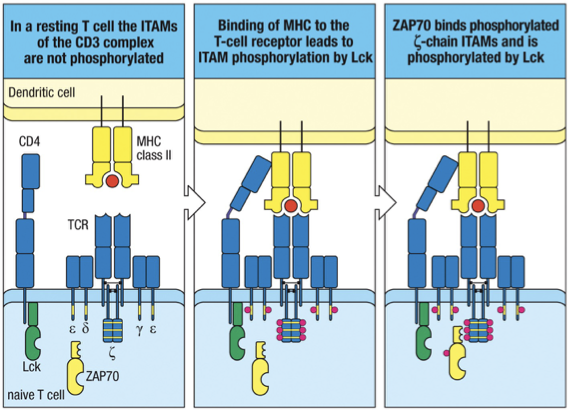
Successful recognition of antigen leads to clustering of TCR and its co-receptor (CD4 or CD8), which initiates intracellular signaling pathways
This clustering is crucial for signaling, resulting in gene expression changes necessary for T cell activation.
The signal that antigen has bound the TCR is transmitted by the cytoplasmic tails of CD3 chains, associated with the TCR a- & b-chains.
CD3 cytoplasmic tails contain sequences called immunoreceptor tyrosine-based activation motifs (ITAMs), which are the little red dots on the tails of the TCR in the picture.
They associate with cytoplasmic protein tyrosine kinases.
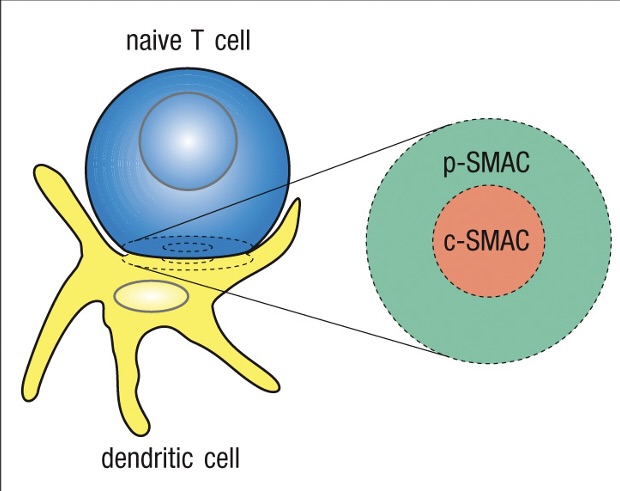
c-SMAC - Central supramolecular activation complex
Clustering of receptors-co-receptors and signaling molecules
p-SMAC - peripheral supramolecular activation complex
Adhesion molecules – forms tight seal
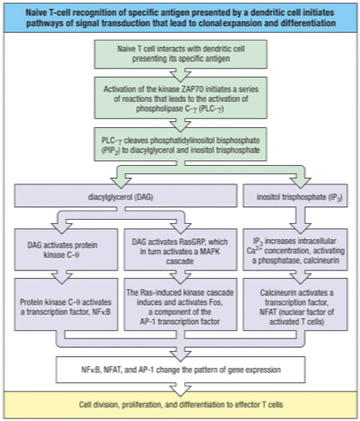
TCR Signaling from Recognition → Effector T Cell
TCR binds peptide–MHC
CD4/CD8 binds MHC → brings Lck close
Lck phosphorylates ITAMs on CD3/ζ chains
ZAP-70 binds phosphorylated ITAMs on CD3/ζ chains→ ZAP-70 is phosphorylated
Activated ZAP-70 → activates PLC-γ
PLC-γ cleaves PIP2 → DAG + IP3
DAG + IP3 → activate transcription factors:
NF-κB
NFAT
AP-1
Transcription factors → drive T cell:
Division (proliferation)
Differentiation → effector T cells
T cell proliferation
Activation of T cell by professional APC initiates a program of differentiation controlled by the cytokine IL-2.
A burst cell division
Acquisition of effector function
IL-2 is synthesized and secreted by activated T cells.
IL-2 binds IL-2 receptors of T cell to drive clonal expansion of the activated cell.
IL-2 is crucial for this activation; without it, naive T cells would not undergo differentiation.
IL-2 production requires both the signal delivered through the TCR:co-receptor complex binding to peptide:MHC and the co-stimulatory signal delivered through CD28 binding to B7.
Signals through the TCR:CD3 complex activate NFAT activates transcription of the IL-2 gene (IL-2 mRNA is inherently unstable).
Co-stimulation signal (CD28 on T cell : B7 on APC) stabilizes the IL-2 mRNA which causes an increase in the synthesis of IL-2 by T cells (20-30 times).
Co-stimulation also activates other transcription factors that stimulate the transcription of the IL-2 gene (3 times).
Principal effect of co-stimulation is to increase synthesis of IL-2 by 100 fold
When a TCR on a mature naïve T cell binds to a peptide:MHC complex on an APC that does not express the co-stimulatory molecule B7 the T cell becomes nonresponsive or anergic and cannot be activated with subsequent antigen encounter.
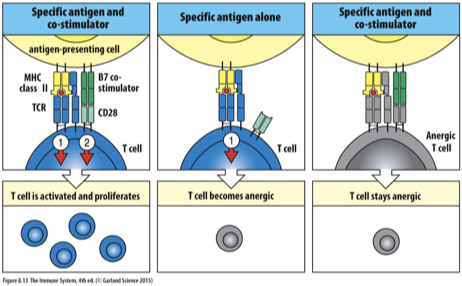
Some mature naïve T cells may be specific for self-proteins expressed by cells not found in the thymus.
These T cells will not be activated because the cells expressing these self antigens will not express the co-stimulatory molecule, B7.
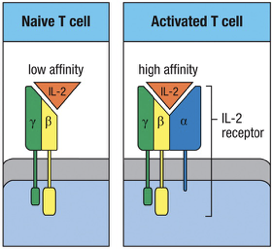
IL-2 Receptor on T cell
Naïve T cells express the low-affinity receptor for IL-2 which consists of and chains.
Activation of a naïve T cell by the recognition of a peptide:MHC complex accompanied by co-stimulation induces the synthesis and secretion of IL-2 and the synthesis of the IL-2 receptor chain.
chain combines with the and chains to make a high-affinity receptor for IL-2.
IL-2 binds to the IL-2 receptor producing an intracellular signal that promotes T-cell proliferation.
IL-2 Importance
Prevention of host vs. graft rejection
Cyclosporin A and tacrolimus – disrupts the production of TCR signals
Rapamycin – disrupts signals from the IL-2 receptor
Prevents adaptive immune response against the foreign MHC peptide complexes on the donor tissue.
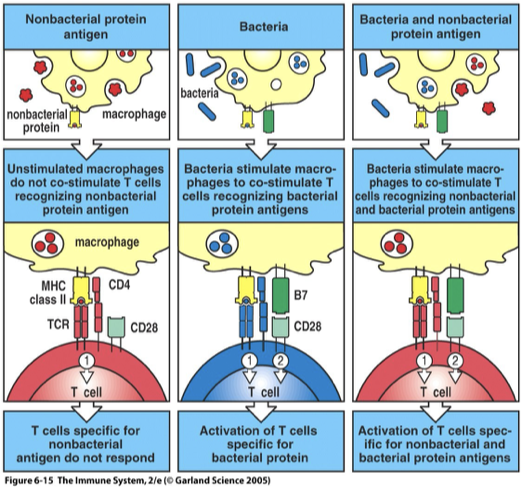
NonBacterial Antigen vs Bacterial Antigen
Immunization of protein antigen alone rarely induces an immune response.
Protein antigens + certain bacterial breakdown products (adjuvants) are required for a strong immune response.
Microbial products, known as adjuvants, induce co-stimulatory activity in P-APC’s.
Whole microorganism = more effective vaccines than highly purified Ag molecules
Mechanism which allows the immune system to distinguish between antigens borne by infections agent and antigens associated with innocuous proteins.
Nonbacterial protein antigens
Often lack PAMPs
→ failure to stimulate APC (macrophage)
→ no B7 (Signal 2)
Only Signal 1 → no activation
Effector T Cells
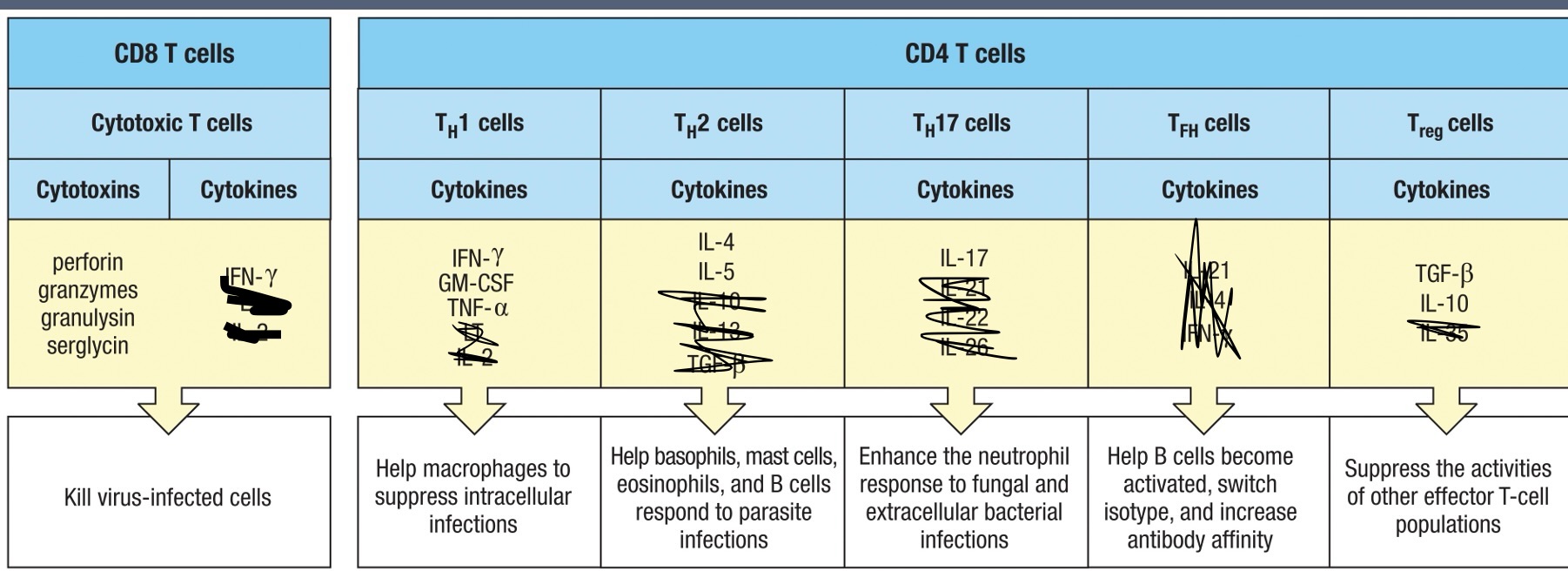
CD8 T Cell Mechanism of Action
Descriptive process of how CD8 T cells induce apoptosis in infected cells:
Recognition of antigens presented on MHC class I molecules leads to the release of cytotoxic granules (perforins and granzymes).
Perforins create pores in target cells.
Granzymes enter through these pores to activate apoptosis mechanisms (not to cause lysis).
Granulysin aids granzymes
Serglycin holds the perforin, granzymes, and granulysin together to form a complex that ensures their efficient delivery to the target cell
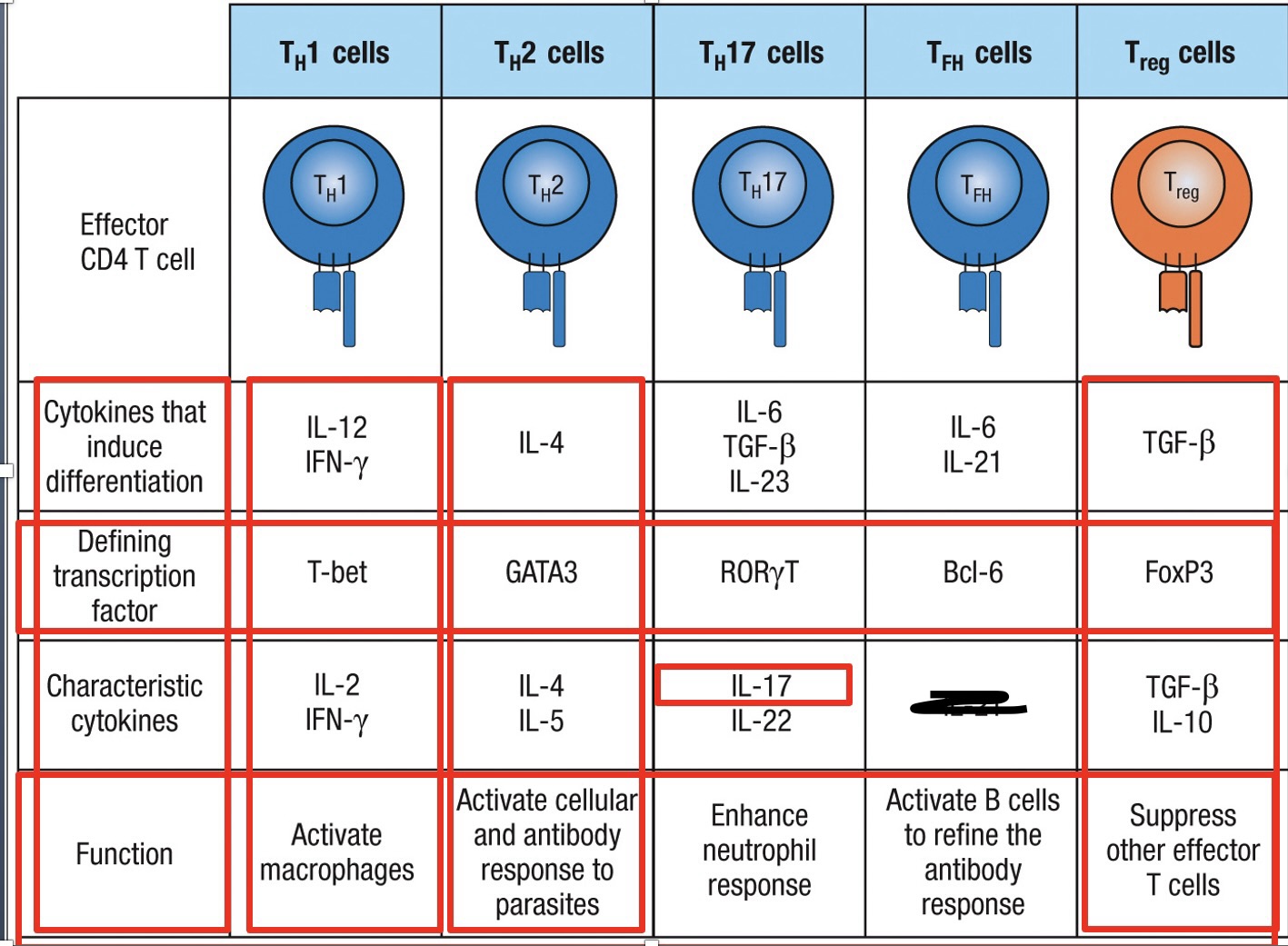
Effector CD4 T cells
TH1 – increases inflammation at the site of infection
Main defense against intracellular viral and bacterial infections
Activated by IL-12 (dendritic cells and macrophages) and IFN-y secreted by NK cells during innate immune response
Activation of transcripion factor T-bet differentiation into TH1 effector
TH2 – defense against parasites in tissues and surfaces
Does not lead to inflammation
Induce IgE production from B-cells
Bind to mast cells, basophils and activated eosinophils
Activated by IL-4 environment, this leads to activation of transcription factor GATA3 differentiation into TH2 effector cell
TH17 – recruits neutrophils to site of infection
Secrete IL-17 which induces production of CXCL8 from epithelial cells and stromal cells
TGF-b and IL-21 or IL-6 activate transcription factor RORyT differentiation into TH17 effector
TFH – Help naïve B-cells in follicles/germinal centers become plasma cells or memory cells
Expresses CXCR5 binds to CXCL13
TFH travels to follicle
B cell isotype switching
IL-6 activates transcription factor Bcl6 TFH effector
Treg – primary cells involved in regulating the immune reponse.
Interact with cells to down regulate their activity
In presence of TGFb the naïve T cell activates transcription factor FoxP3 Treg cell
Produce IL-10 and TGFb (Reduce inflammation and immune response)
Natural (nTreg) and induced (iTreg) T regulatory cells.
nTreg – differentiate in thymus
iTreg – differentiate in periphery
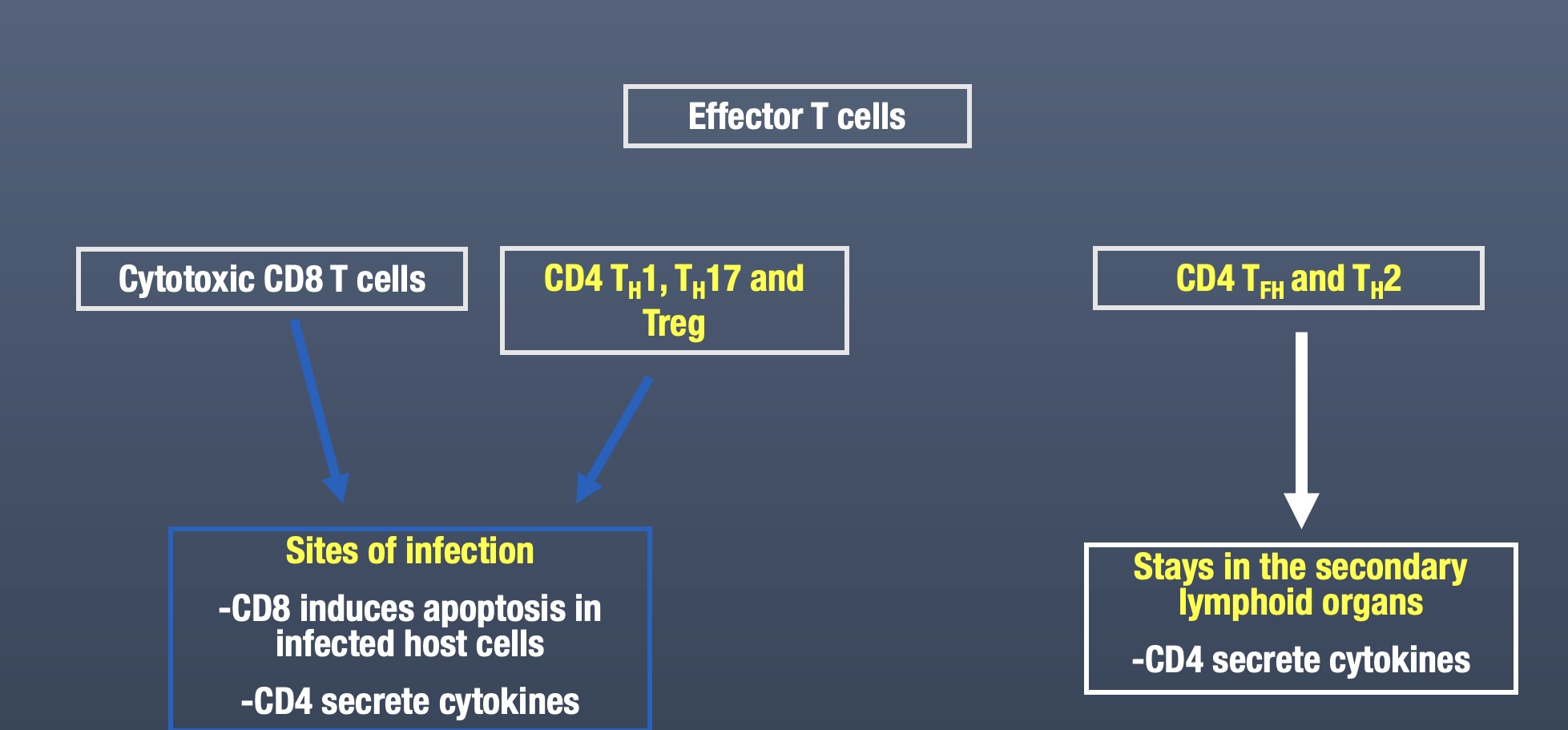
TH1 vs TH2 response
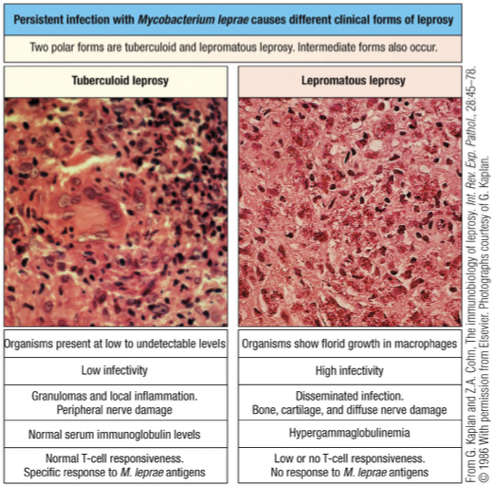
Tuberculoid Leprosy - strong TH1 response (effective); IL2 and IFN-y cytokines found
Lepromatous leprosy - weak TH2 response (ineffective); IL4 and IL5 cytokines found
Simplified
If Th2 dominates during a bacterial or viral infection (non-parasite), it’s “useless” and can impair proper immunity.
Why: Th2 mainly drives antibody responses and eosinophils, which do not effectively clear intracellular bacteria or viruses.
What should happen: Th1 cytokines (IFN-γ, IL-2) activate macrophages and cytotoxic T cells to kill infected cells.
CD8 T cells
The activation of naïve CD8 T cells requires stronger co-stimulation than CD4 T cells.
CD8 T cells stimulated by antigen and co-stimulation will synthesize IL-2 and the high affinity IL-2 receptor, which induces their proliferation and differentiation.
Under conditions of suboptimal co-stimulation, CD4 T cells can help activated naïve CD8 T cells.
CD4 and CD8 T cells must recognize their specific antigen on the same APC, then the CD4 T cells will secrete IL-2 which can then bind to the IL-2 high affinity receptor and activate the CD8 T cell.
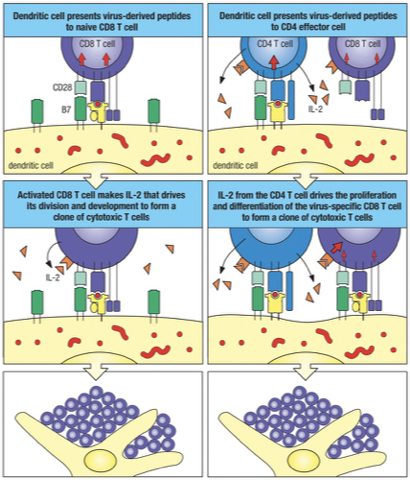
A major change is that…
Effector T cells can respond to their specific antigen without the need for co-stimulation via B7-CD28 interaction.
Effector T cells can respond to antigen presented on cells other than professional antigen-presenting cells.
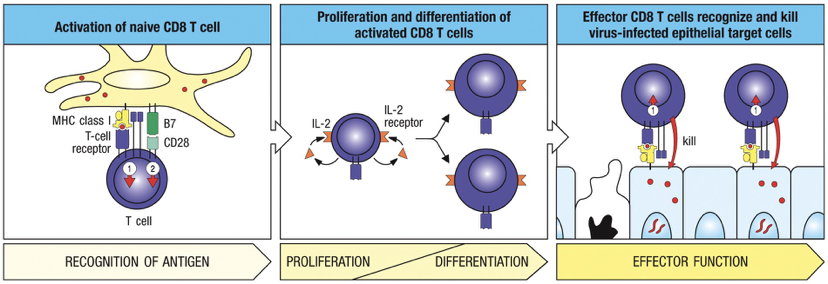
Once licensed by dendritic cell (with B7: CD28), anything that has same MHC I combination does not need activation; CD8 T cell is licensed to kill
Adhesion Molecules: Naïve VERSUS Effector
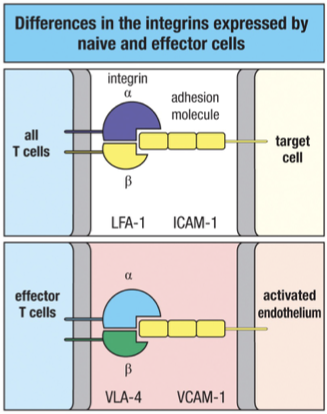
Naïve = Entering Node
L-selectin (selectin, on T cell → binds addressins on HEV) → lymph node homing
LFA-1 (low) (integrin, on T cell → binds ICAM-1 on APC/endothelium) → weak adhesion
CD2 (low) (Ig superfamily, on T cell → binds LFA-3 on APC/target) → weak stabilization
Effector = Sticky + Site
❌ No L-selectin (can’t home to lymph nodes if inside binding to APC)
LFA-1 (high) (integrin, on T cell → binds ICAM-1 on target) → strong adhesion (after TCR activation)
CD2 (high) (Ig superfamily, on T cell → binds LFA-3 on target) → stabilizes binding
VLA-4 (integrin, on T cell → binds VCAM-1 on inflamed endothelium) → homing to infection site
Cytokines VERSUS Cytotoxins
Cytokines, which alter the behavior of target cells.
All effector T cells produce cytokines (just different types and in different combinations).
Cytotoxins, secreted cytotoxic proteins used to kill target cells.
Cytotoxins are specialized products of cytotoxic CD8 T cells.
Secreted Cytokines
Secreted cytokines and related membrane-bound proteins that act through cell-surface receptors generally to induce changes in gene expression within target cells.
Work locally and work over short period of time
Autocrines: act on the cell that produced them
Paracrines: act locally on another cell
Colony stimulating factors: work at a distance: stimulate bone marrow
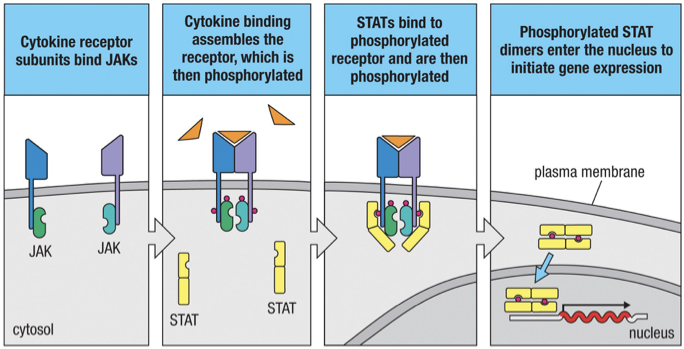
JAK/STAT Cytokine receptor signalling
The cytoplasmic tail of most cytokine receptors are associated with protein kinases called Janus kinases (JAKs).
Cytokine binding causes dimerization of the cytokine receptors which stimulate the JAKs to phosphorylate members of a protein family called STATs (Signal Transducers and Activators of Transcription).
On phosphorylation, two STAT molecules dimerize and move from the cytoplasm to the nucleus where they activate specific genes.
The genes that are activated is determined by the cytokine/s that were bound.
Cytotoxic CD8 T Cells
Function of cytotoxic CD8 T cells is to kill cells that have become overwhelmed by intracellular infection.
Infected cell is sacrificed to prevent spread of infection to healthy cells.
Delivery of Cytotoxins
Lytic granules are modified lysosomes with a mixture of cytotoxins.
CD8 T cells synthesize cytotoxins in inactive forms and package them into lytic granules when T cells are activated by specific antigen in the secondary lymphoid organs.
CD8 T cells then migrate to sites of infection and will recognize specific peptides (made from proteins from infected cell) in the context of MHC class I molecules presented by the infected cell.
TCR binding signals the cytotoxic T cell to secrete the contents of lytic granules directly onto a small localized area on the surface of the infected target cell.
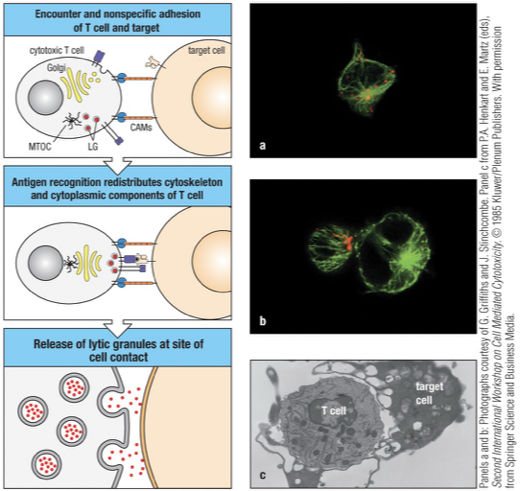
LG= lytic granules (red)
MTOC= microtubule organizing center (microtubules – green)
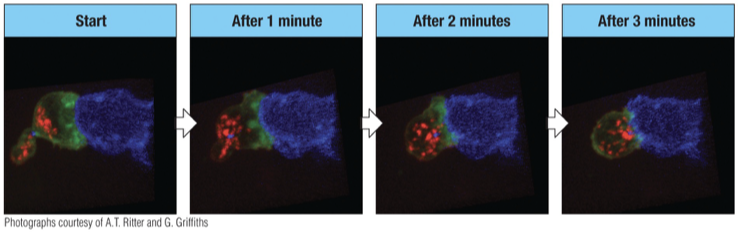
Contact is made with target cell and granules are distant from the point of contact.
After one minute, granules have begun to move toward the target cell.
After 3 minutes granules are almost ready to be released.
Selective Killing
Specific recognition of peptide:MHC complex on an infected cell by a cytotoxic CD8 T cell (CTL) programs the infected cell to die and leaves healthy cells alone.
The CTL detaches from the first target cell (which dies), synthesizes a new set of lytic granules and then seeks out and kills another infected cell (new target).
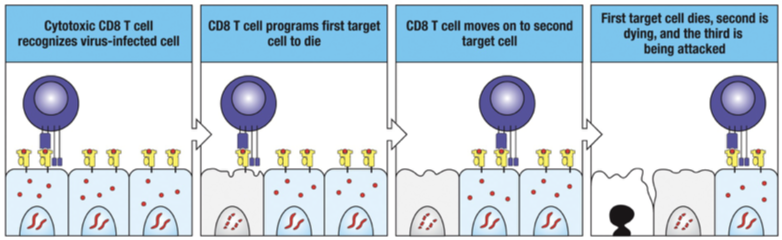
CD8 contributing cytokines
Secretion of interferon-gamma (IFN-y) inhibits the replication of viruses in the infected cells
IFN-y also increases the processing and presentation of viral antigen by MHC class I molecules.
IFN-y activates macrophages in the vicinity of the cytotoxic T cells.
Activated macrophages then get rid of dying infected cells helping damaged tissue to repair.
Apoptosis VERSUS Necrosis
Necrosis = death due physical or chemical injury in which cells lyse and disintegrate.
Apoptosis or programmed cell death = cell suicide in which cells shrivel or shrink but remain intact.
Apoptosis of the target cell is induced by the cytotoxic CD8 T cells releasing cytotoxins.
Target cells die by apoptosis which prevents pathogen replication as well as the release of the infectious pathogen.
Apoptosis
Apoptotic cell in late stages has very condensed nucleus, no mitochondria, and the cytoplasm and cell membranes are largely lost through vesicle shedding.
plasma membrane still defined
Necrosis
cell lysis and leakage of contents
plasma membrane poorly defined
TH1 CD4 cells induce macrophages to become activated
Macrophage have receptors that bind to microorganisms and facilitate their phagocytosis, destruction and intracellular degradation.
As a result, macrophages present pathogen-derived peptide on MHC class II molecules to activate effector TH1 cells.
Thus, a principal function of TH1 cells is to activate macrophages to:
Increase their phagocytic ability.
Increase their capacity to kill ingested microorganisms.
Activation Signals
Macrophages require two signals for activation, both of which are delivered by TH1 cells.
The primary signal is provided by IFN-y, the characteristic cytokine produced by TH1 cells.
The second signal is delivered by the CD40 ligand (CD40L) on T cells interacting with the CD40 receptor on macrophages.
Together these newly synthesized proteins activate the macrophage to kill the bacterial living inside of its vesicles.
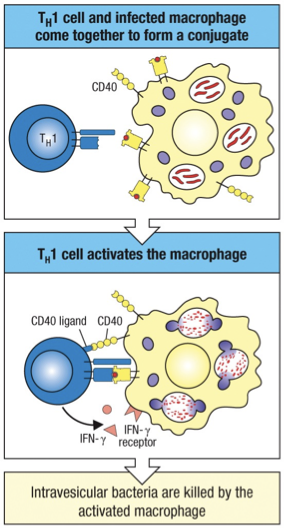
Essentially; TH1 cells coordinate the host response to intravesicular pathogens
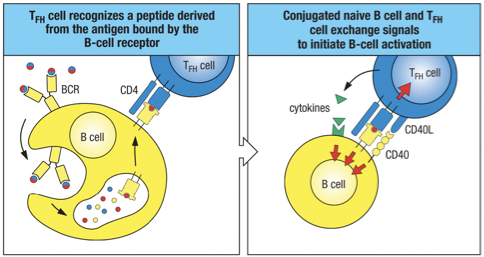
TFH/B Cell Interaction
The specific interaction of an antigen binding B cell with a TFH cell leads to the expression of CD40 ligand (CD40L) on the T cell; also known as CONJUGATE PAIRS
For B cell to receive T cell help, both cells must recognize the same antigen – cognate interactions.
Linked recognition – B cell may recognize a surface epitope, but the peptide presented to the T cell may not be the same exact epitope (remember it will be a peptide).
Regulatory T cell Inhibition (T)
Regulatory or suppressor CD4 T cells make inhibitory cytokines.
IL-4, IL-10, TGF-B with high expression of CD25 ( chain of IL-2 receptor)
Inhibition depends on physical contact between the regulatory CD4 T cell and its target
FoxP3 Transcription Factor