Sensory Reception
Sensory systems include vision, somatic sensory/touch, gustatory/taste, olfaction/smell, auditory/hearing, balance/movement and proprioception. Proprioception is the ability to know where your different body parts are in relation to other things.
Sensory reception is needed to maintain homeostasis, detect and react to changes in environment, and protect the body from noxious stimuli such as extreme heat.
stimulus → sensory receptor → afferent neuron → CNS → efferent neuron(s) → effector organ (muscle or gland) → response
Receptors which respond to chemical pain, temperature (thermal pain), pressure (mechanical pain) or touch are known as cutaneous receptors. Receptors which detect body position are known as proprioceptors. There are specialised receptors for sight, hearing, taste and smell.
Chemical pain stimulates unmyelinated C fibres, as does thermal pain and mechanical pain. Thermal and mechanical pain also stimulate myelinated A-delta fibres. Touch stimulates myelinated A-beta fibres. Changes in body position stimulates myelinated A-alpha fibres. C fibres have the slowest rate of transmission of 0.5 to 2 m/s whereas A-alpha fibres have the fastest rate of transmission of 80-120 m/s.
Pain receptors are activated by the release of ATP and prostaglandins from damaged or stressed cells ATP binds purinergic receptors allowing sodium ions to enter the cell and prostaglandins bind prostaglandin receptors. Na+ accumulates leading to an action potential sent via the spinal cord.
In 2019, a paper was published about specialised Schwann cells which form a mesh in the skin and associate with C-fibres. The cells were able to detect mechanical pain and transmit signals to C-fibres.
Different receptors are activated in extreme cold (<15 degrees, TRPM8) and extreme heat (>45 degrees, receptor TRPV1). Menthol also activates TRPM8 and capsaicin (chillies) activates TRPV1.
Pacinian corpuscles in the skin are sensitive to pressure and vibration. They consist of a nerve ending surrounded by layers of highly specialised connective tissue. The nerve ending contains specialised stretch mediated sodium ion channels.

Other sensory receptors in the skin include Merkel’s discs, Messner’s corpuscles and Ruffini’s endings. They are sensitive to various types of touch. For example, Merkel discs detect texture and sharp edges. Physical stimulation of Merkel cells leads to release of %-HT, activating 5HT3 and 5HT2 receptors on A-beta fibres leading to depolarisation. Merkel cells also protect against innocuous mechanical itch via stretch-activated Piezo2 channels. Reduced Merkel cells in old age or dry skin cause decreased light touch sensing and increased mechanical itch.
Receptors for pain, temperature and body position are slowly adapting, allowing for continual awareness and response. Some mechanoreceptors are rapidly adapting so that ‘unimportant’ information can be ignored.
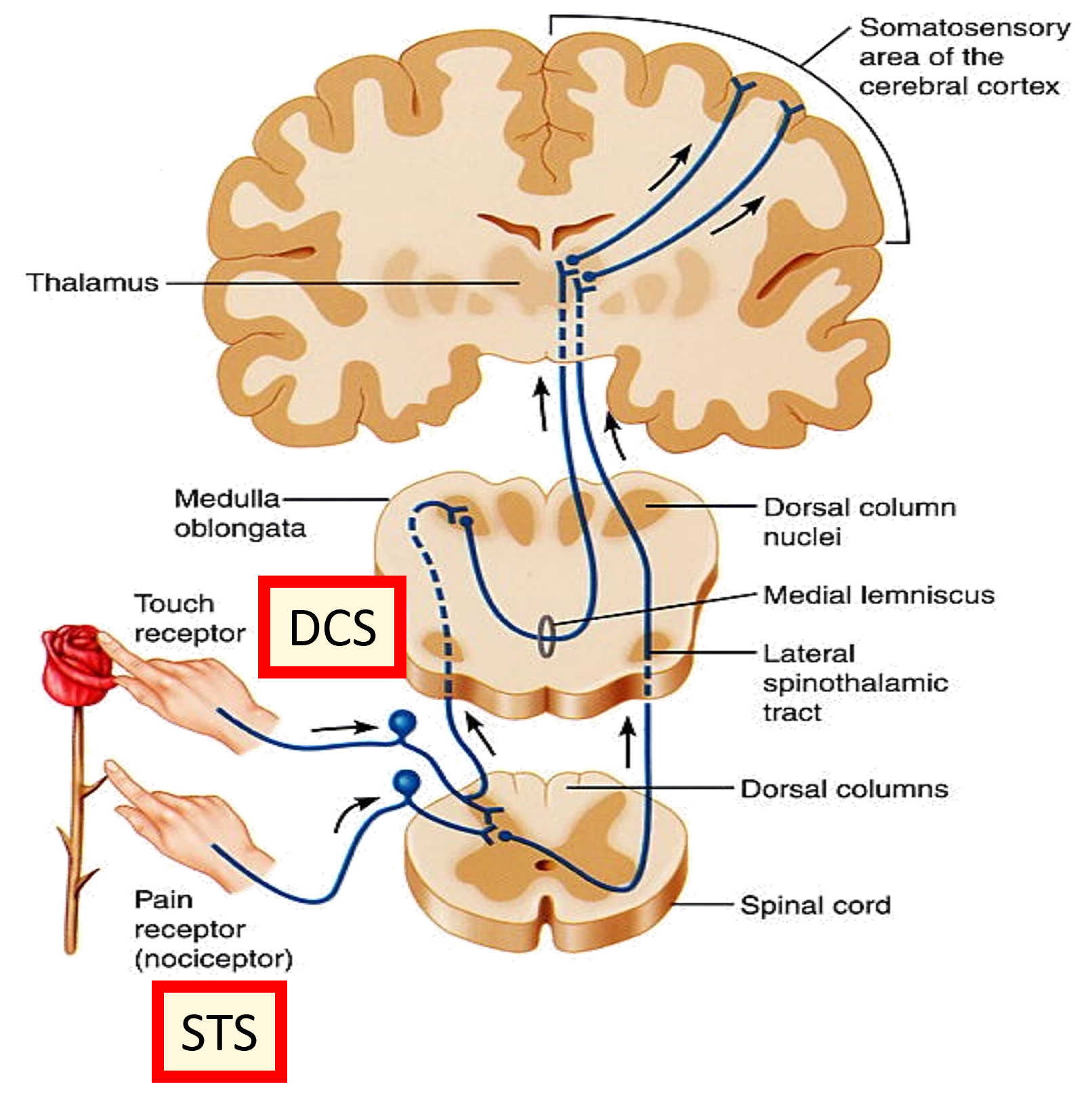
The dorsal column system responds to fine touch, pressure and proprioception. The spinothalamic system responds to pain, temperature and coarse touch. Decussation takes place, where the pathways cross to the opposite side from which the stimulus was experienced. This means sensory information is processed on the opposite site of the brain. Sensations which are to be consciously experienced are sent to the post-central gyrus via 3rd order neurons.
Number of neurons controlling each area of the body varies massively. The largest proportions control the hands and lips/face etc.
The two types of nociceptor are A-delta and C fibres. A-delta are myelinated and transmit sensations of fast, sharp pain. C fibres are unmyelinated and transmit sensations of slow, dull aching pain. Polymodal nociceptors are activated by high intensity mechanical, chemical and thermal stimuli and the fibres are C fibres.
Stimuli activate nociceptors which send action potentials down afferent neurons to the spinal cord. At the pre-synaptic axon terminal, substance P is released from the afferent neuron and a new action potential is created in a second order neuron. This is the spinothalamic pathway.
Referred pain is where pain is received to come from a different part of the body from where the pain signals are generated eg someone experiencing a heart attack may feel pain/discomfort in the arm, back or neck region. This is because the sensory receptors enter the same neuropathways.
Interneurons release enkephalins on either descending stimuli from the CNS or localised 5_HT to activate pre- and post-synaptic opiate receptors at the spinal sensory synapse. This causes GPCRs to couple to Gi leading to decreased cAMP → decreased PKA, modulating K+ release and decreasing neuronal excitability in C fibres as a result.
The gate control theory of pain modulation suggests that the spinal cord contains a neuronal ‘gate’ which either blocks pain signals or allows them to continue to the brain. Signals sent along C fibres alone are not inhibited so a strong signal is sent to the thalamus. However action potentials sent along A-beta fibres (dorsal columns) may depolarise inhibitor interneurons, preventing as many signals from being sent along the relay neurones to the thalamus and resulting in a smaller/no sensation of pain.
A-delta fibres synapse in a different area of the spinal cord to C fibres and there are no interneurons that release enkephalins. No opiate receptors present in the area means morphine is ineffective. As a result, sharp pain is harder to treat.
Everybody perceives pain differently due to prior experience, expectation of pain, mood eg anxiety/depression, genetics etc.
Pain can be treated with non-steroidal anti-inflammatory drugs eg aspirin and ibuprofen which block COX-1 and COX-2, therefore blocking inflammation. Noxious pain signals can be transmitted via prostaglandins.
Local anaesthetics such as lidocaine block voltage-gated sodium ion channels in the area they’re injected into, blocking action potentials.
Opioids such as morphine are highly effective pain relief and work at a few different sites. Morphine activates descending inhibition, inhibits transmitter release, hyperpolarises neurons and potentially inhibits discharge of primary afferents.
Acute pain is beneficial to us as it helps to keep us safe, however chronic pain is not beneficial. Neuropathic pain occurs as a result of nociceptor pathways changing leading to neuronal death.