lecture 4: MHC
Mind Map: Major Histocompatibility Complex (MHC)
Central Idea: Major Histocompatibility Complex (MHC) is a group of genes that play a crucial role in the immune system.
Main Branches:
MHC Structure
MHC Classes
MHC Functions
MHC Structure:
MHC I
Alpha chain
Beta-2 microglobulin
MHC II
Alpha chain
Beta chain
MHC Classes:
MHC I - intracellular pathogens
Present on all nucleated cells
Present antigens to CD8+ T cells
MHC II - extracellular pathogens that have been bought in
Present on antigen-presenting cells (APCs)
Present antigens to CD4+ T cells
MHC Functions:
Antigen Presentation
MHC I pathway
MHC II pathway
Immune Response Regulation
Self-recognition
Immune tolerance
Transplant Compatibility
MHC matching
Graft rejection
MHC I Pathway:
Antigen processing
Proteasome degradation
Transporter associated with antigen processing (TAP)
Antigen presentation
Peptide binding to MHC I
Presentation on cell surface
MHC II Pathway:
Antigen processing
Endocytosis or phagocytosis
Lysosomal degradation
Antigen presentation
Peptide binding to MHC II
Presentation on APC surface
Self-Recognition:
MHC restriction
Positive selection in thymus
Immune Tolerance:
Negative selection in thymus
Central and peripheral tolerance mechanisms
MHC Matching:
Importance in organ transplantation
HLA typing for compatibility assessment
Graft Rejection:
Recognition of non-self MHC
Activation of immune response
GENERATION OF T CELL
There are two major intracellular compartments separated by membranes. the cytosol and the vesicular system. Viruses can directly penetrate into the intracellular compartment by entering through membranes, while the extracellular compartment is what has been bought into the cell through endosomes. This means that information and material can be bought into the cell
The MHC class 1 and 2 peptides deliver peptides to the cell surface from these two intracellular compartments. Cytosolic pathogen peptides bind to MHC 1 and are presented to CD8+ T cells leading to cell death. endocytic (both intravesicular and extracellular) pathogen peptides bind to MHC 2 and are presented to CD4+ T cells. For intravesicular pathogens this activates to kill intravesicular bacteria while for extracellular pathogens, the B cells are activated to secrete Ig to eliminate them.
B cells need to be able to bring the antigen into the cell, break it down and present it by MHC 2 to T cell which can activate it to secrete Ig
MHC class 1
peptides that bind to class 1 are actively transported from the cytosol to the endoplasmic reticulum via the TAP complex. Peptides can be from pathogens or degraded self proteins.
Tap is heterodimer that makes a pore in the ER membrane where MHC 1 is being made. This allows it to be loaded onto the MHC. TAP is varied among people and different variations may allow you to be more likely to get a specific disease or infection.
Peptides for transport into the ER are generated in the cytosol by proteolytic digestion in the proteasome. Before peptides can go through the TAP complex, they have to be broken down in order to do this. This happens through the proteasome.
Structurally, It has a proteasome activator domain at the top and bottom, then an alpha domain and then a beta one which contains a catalytic chamber. Proteins that need to be degraded gets flagged by ubiquitination which then allows it to go into the proteasome to break down into small chunks. When infected, the proteasome beta unit changes slightly so that when the protein is being broken down, the C terminal will have a hydrophobic amino acid which will guide it into the MHC molecule.
Newly synthesised MHC 1 molecules are retained in the endoplasmic reticulum until they bind a peptide. The MHC itself is a singular polypeptide that folds into three domains and has a beta 2 microglobulin added to it. Calnexin stabilises it until this binds. After, a complex of chaperone proteins (calreticulin and ERp57) bind to TAP by tapasin. MHC 1 is cotranslationally inserted into the ER membrane. Cytosolic proteins and DRiPs (defective ribosomal products) are degraded to peptide fragments by the proteasomes. TAP delivers peptides to the ER. A peptide binds the MHC 1 molecule which completes its folding. The MHC id released from the TAP complex and is exported to the cell membrane. The associated chaperones fall off. CD8+ T cells can now recognise the antigen and mount an immune response
Some viruses inhibit this virus such as adenovirus which produces E19 that competes with tapasin. This inhibits peptide loading onto the MHC.
MHC Class 2
MHC 2 is also made on ribosomes on the ER and cotranslationally inserted onto the ER membrane. To prevent loading of cytosolic proteins, the invariant chain physically blocks the binding groove as it is being made. This prevents any peptides in the ER binding. As MHC is travelling towards the acidified vesicle, the end parts of the invariant chain starts breaking down due to the acid, until there is the CLIP part of the invariant chain left.
The antigen is taken up from the extracellular space into intracellular vesicles. In the early endosomes at neutral pH, the proteases are inactive however after acidification of the vesicles, the proteases degrade antigens into peptide fragments, The vesicles then fuse with other vesicles containing the MHC 2 molecules. If the pathogenic material matches with the anchor residue pattern, it is then put onto the cell surface
Endocytosed antigens are degraded to peptides in the endosomes but CLIP blocks the binding of the peptide to MHC 1. HLA-DM binds to MHC 2 and releases CLIP which allows the antigenic peptide to bind. MHC 2 can then travel to the cell surface. CD4+ T cells can now come recognise the antigen and mount an immune response
Sometimes, extracellular peptides need to be loaded onto MHC 1 in order to activate CD8+ T cells. This is so they can differentiate into Tc cells. This is stimulated by MHC 1 but extracellular peptides are normally loaded onto MHC 2. For this to happen, cross presentation occurs.
Bits of particle could be on the outside of the infected cell and bought in through the endosomal pathway. This can then be loaded into the MHC 1 pathway by translocation into the cytosol however, the exact mechanism is unknown.
MHC 2 molecules can also present cytosolic proteins by autophagy. Autophagy is the natural process where some damaged organelles and cytosolic proteins are delivered the autophagosomes from proteolytic breakdown. These peptides can then be expressed on MHC 2.
In essensence, there is cross presentation to act as a safety check to make sure there is the right response and if the antigen doesn’t end up in the right compartment, there is a way to fix it.
GENETICS
Certain pathogens need a T helper response rather than a cytotoxic T cell so it is very important that the right bit of antigenic information is in the right presentation pathway. This means that naive T cells can learn what they are responding to and differentiate accordingly
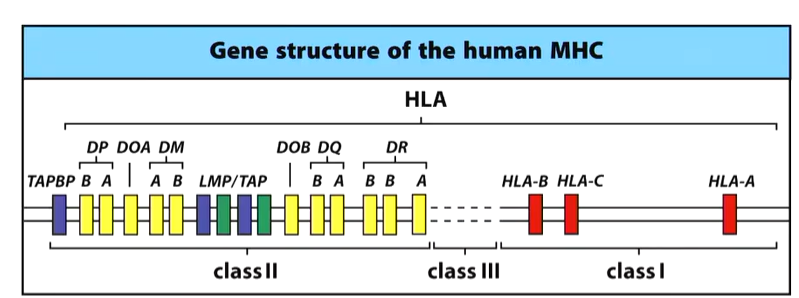
Chromosome 6 contains the genetic information for the MHC receptors. Class 1,2 and 3 genes are located in clusters along this chromosome. These act as HLA loci which codes for the polypeptide chain that folds into the three domains for MHC.
On this region of DNA, there are three alleles (HLA1/2/3) which make a different MHC 1 polypeptide chain. In the class 2 zone, there are associated genes. MHC2 has two polypeptide chains which re represented by DP a and b. DQ and and b and DR a,b,b are the alleles responsible for the actual MHC 2 molecule. TAPBP encdoes tapasin but has some role in MHC 2 but puts MHC 1 together. DOA and DOB play a role in the production of the DM molecule that allows CLIP to detach. LMP is part of the proteasome process that allows proteins to come in through the MHC 1 pathway.
HLA stands for human leukocyte antigen which is system of genes that encode for cell surface proteins needed in the immune system such as MHCs.
The protein products of MHC class 1 and 2 genes are highly polymorphic. This means that MHC can be both polymorphic and polygenic. You inherit all 6 HLAs
different genes = polygenic
different versions of the genes = polymorphic
This means you van have lots of diversity in MHC 1
For MHC 2, it is the same but with DP a/b, DQ a/b and DR a/b/b which are also inherited as a haplotype
This is important for transplant sites as you only share a similar pattern of MHC 1 to someone very closely related. This variation is important as it allows variability in the binding groove. This is important for how the immune cells recognise MHC molecules. MHC polymorphism affects antigen recognition by T cells by influencing both peptide binding and the contacts between T cell receptor and MHC molecules
T cells binding too tightly to MHC is bad because if it binds more strongly to the antigen, it makes a massive immune response to anything. An example of this is toxic shock syndrome where the toxin binds to MHC and TCRs through cross binding. This means that there is no specificity so T cells think they’ve gone through every stage. It needs to bind just right