B3.1 - Gas exchange
Standard level
3.1.1 Gas exchange as a vital function in all organisms
Gas exchange is a vital function in all living organisms as it facilitates the interchange of materials for metabolic reactions.
Oxygen is required for aerobic respiration, whereby cells produce large amounts of chemical energy (ATP)
Carbon dioxide is a waste product of this process and excess levels must be removed from the organism
In animals, respiratory gases are transported within the bloodstream and gas exchange is mediated by specialised ventilation systems (e.g. lungs, gills, etc.)
In larger organisms, the distance from the centre of the organism to the exterior is greater – meaning gas exchange would not be efficient without a means of continuous ventilation
Larger organisms also have a smaller SA:Vol ratio – this makes necessary a specialised ventilation system to ensure there is adequate gas supply to the tissues
Different types of organisms have evolved distinct systems for gas exchange that are suited to their environment – the complexity of these systems is typically correlated to organism size ADD MORE ABOUT FISH AND INSECTS
For small multicellular organisms (such as flatworms), simple diffusion across the external surface is sufficient to meet their respiratory needs
Insects have a respiratory system that is independent of the circulatory system – instead they have a tracheal system to move gases throughout the organism
Aquatic organisms (such as fish) have evolved heavily folded structures called gills to take up the dissolved oxygen from within the water
In mammals, ventilation is mediated by spongy respiratory structures called lungs, which exchange gases between the bloodstream and the atmosphere
3.1.2 Properties of gas exchange surfaces
Ventilation surfaces must possess certain properties to allow for adequate amounts of gas exchange:
Surface area: A large surface are will allow sufficient concentrations of gases to be exchanged
Moisture: It is easier for gases to diffuse across membranes when they are dissolved in a solution
Absorptive: Surfaces must be adequately permeable to enable the movement of respiratory gases
Rich blood network: Gas exchange surfaces must be connected to a dense capillary network
Thin tissue layer: Ventilation surfaces are lined by a single layer epithelium to optimise gas exchange
Alveoli are the site of gas exchange within the lungs and are composed of two types of alveolar cells (pneumocytes)
Type I pneumocytes are flattened and extremely thin – they function as the gas exchange surface
Type II pneumocytes are responsible for the secretion of pulmonary surfactant, which reduces surface tension in the alveoli
Pulmonary surfactant prevent the sides of the alveolus adhering to each other by reducing surface tension
As an alveolus expands with gas intake, the surfactant becomes more spread out, increasing surface tension and slowing the rate of expansion
This ensures that all alveoli inflate at roughly the same rate and to proportionate dimensions – optimising gas exchange (o2 in, co2 out)
3.1.3 Maintenance of concentration gradients at exchange surfaces in animals
Because gas exchange is a passive process, a ventilation system is needed to maintain a concentration gradient at exchange surfaces
Oxygen is constantly being consumed by cells for aerobic respiration, while carbon dioxide is continually being released
Ventilation systems function to continually cycle fresh gases into the tissues from the atmosphere (terrestrial animals) or water (aquatic animals)
This means oxygen levels stay high within the respiring tissues and carbon dioxide levels remain low
Respiratory gases are transported within the bloodstream, meaning that ventilation systems need to be connected to a dense network of blood vessels with continuous blood flow
Ventilation systems are also structured to have a very large surface area, so as to increase the overall rate of gas exchange
3.1.4 Adaptations of mammalian lungs for gas exchange
In mammals, gas exchange involves a specialised structure called the lungs (responsible for pulmonary ventilation)
Air enters the respiratory system through the nose or mouth and passes through the pharynx to the trachea
(The trachea is ciliated and lined with mucus in order to trap and expel any foreign particulate matter)
The air travels down the trachea until it divides into two bronchi (singular: bronchus) which connect to the lungs
(The right lung is composed of three lobes, while the left lung is only comprised of two, its smaller due to position of heart)
Inside each lung, the bronchi divide into many smaller airways called bronchioles, greatly increasing surface area
The bronchioles contain smooth muscle innervated by the autonomic nervous system to allow for the regulation of air flow
Each bronchiole terminates with a cluster of air sacs called alveoli, where gas exchange with the bloodstream occurs
Certain alveolar cells secrete pulmonary surfactant – this reduces surface tension and prevents alveoli from collapsing
The alveoli are connected to a dense network of capillary beds – this allows for the optimised exchange of gases with the blood
3.1.5 Ventilation of the lungs
Breathing is the active movement of respiratory muscles that enables the passage of air into and out of the lungs
The contraction of respiratory muscles changes the volume of the thoracic cavity (i.e. the chest)
Changing chest volume creates a pressure differential between the chest and atmosphere – with air then moving to equalise
Respiratory muscles - Because muscles only do work via contraction, different groups of respiratory muscles are required to expand and contract chest volume
Muscles that increase the volume of the chest cause inspiration (breathing in)
Muscles the decrease the volume of the chest cause expiration (breathing out)
The core muscles involved in ventilation are the diaphragm and the intercostal muscles between the ribs (internal and external). Additional accessory muscle groups support the act of breathing – including the abdominal muscles and pectoral muscles.
Breathing - Inspiration (inhaling) and expiration (exhaling) are controlled by two sets of antagonistic (antagonistic pair, when one contracts (shortens), the other one relaxes (lengthens)) muscle groups
The muscles responsible for inspiration are the diaphragm and external intercostals
The diaphragm muscles contract, causing the diaphragm to flatten and increase the volume of the thoracic cavity (chest cavity - space inside your thorax (chest) that contains your heart, lungs and other organs and tissues)
External intercostals contract, pulling ribs upwards and outwards (expanding chest), internal intercostals relax
As the volume of the thoracic cavity increases, the pressure within the lungs decreases below atmospheric pressure and air enters the lungs
The muscles responsible for expiration are the abdominal muscles and internal intercostals
The diaphragm muscles relax, causing the diaphragm to curve upwards and reduce the volume of the thoracic cavity
Internal intercostals contract, allowing the downward and inward movement of the ribcage (deflating the lungs), external intercostals relax
Abdominal muscles contract and push the diaphragm upwards
With the volume of the thoracic cavity reduced, the pressure within the lungs increases above atmospheric pressure and air is expelled from the lungs
3.1.6 Measurement of lung volumes
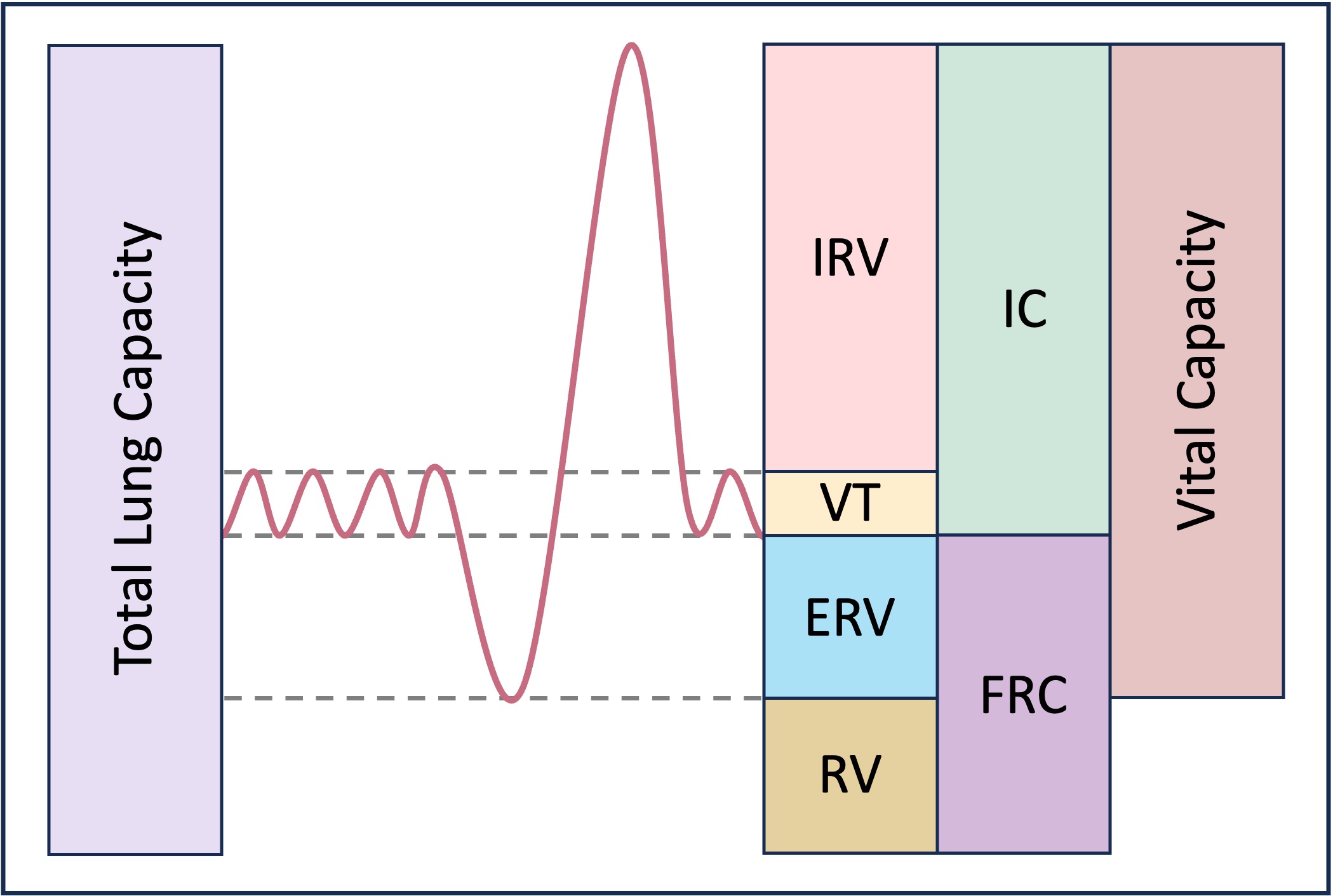
Lung capacity refers to the volumes of air associated with the different phases of the respiratory cycle:
Total lung capacity – Volume of air in the lungs after a maximal inhalation (~ 6 litres in a normal adult male)
Vital capacity – Volume of air that can be exchanged by the lungs via a maximal inhalation and exhalation - VC=TV+IRV+ERV
Residual volume – Volume of air that is always present in the lungs (~ 20% of total lung capacity)
Tidal volume – Volume of air that is exchanged via normal breathing (~ 500 ml per breath)
Inspiratory Reserve Volume – Volume of air able to be inspired with maximum effort after a normal inhalation
Expiratory Reserve Volume – Volume of air able to be expired with maximum effort after a normal exhalation
Inspiratory Capacity – The sum of the inspiratory reserve volume (IRV) and tidal volume (VT)
Functional Residiual Capacity – Sum of the expiratory reserve volume (ERV) and residual volume (RV)
The ventilation rate (breathing frequency) for a typical adult at rest is roughly 12 – 16 breaths per minute
Ventilation rate - number of inhalations or exhalations per minute
(A number of factors contribute to an individual’s total lung capacity and ventilation rate, including height (taller people tend to have larger chests and hence larger total lung capacities), location (people living at high altitudes tend to have larger capacities to compensate for the lower atmospheric pressure)and lifestyle (obese people and smokers tend to have lower capacities and higher rates of ventilation))
Measuring ventilation - Ventilation in humans can be monitored in a number of ways:
Via simple observation (counting number of breaths per minute)
Chest belts and pressure meters (recording the rise and fall of the chest)
Spirometers (recording the volume of gas expelled per breath)
Spirometry involves measuring the amount (volume) and / or speed (flow) at which air can be inhaled or exhaled
A spirometer is a device that detects the changes in ventilation and presents the data on a digital display
3.1.7 Adaptations for gas exchange in leaves
The leaves of a plant play a key role in photosynthesis and function as the site of both light absorption and gas exchange
Leaves are typically broad, flat and thin to maximise their surface area and optimise rates of photosynthesis
The outer surface of the leaf is composed of a thin layer of tissue called the epidermis
A waxy cuticle covers the epidermis to provide an hydrophobic barrier (prevents water loss)
The lower epidermis is has stomatal pores which are openings that allow gas exchange (co2 in, o2 out) and water vapour leaving (transpiration)
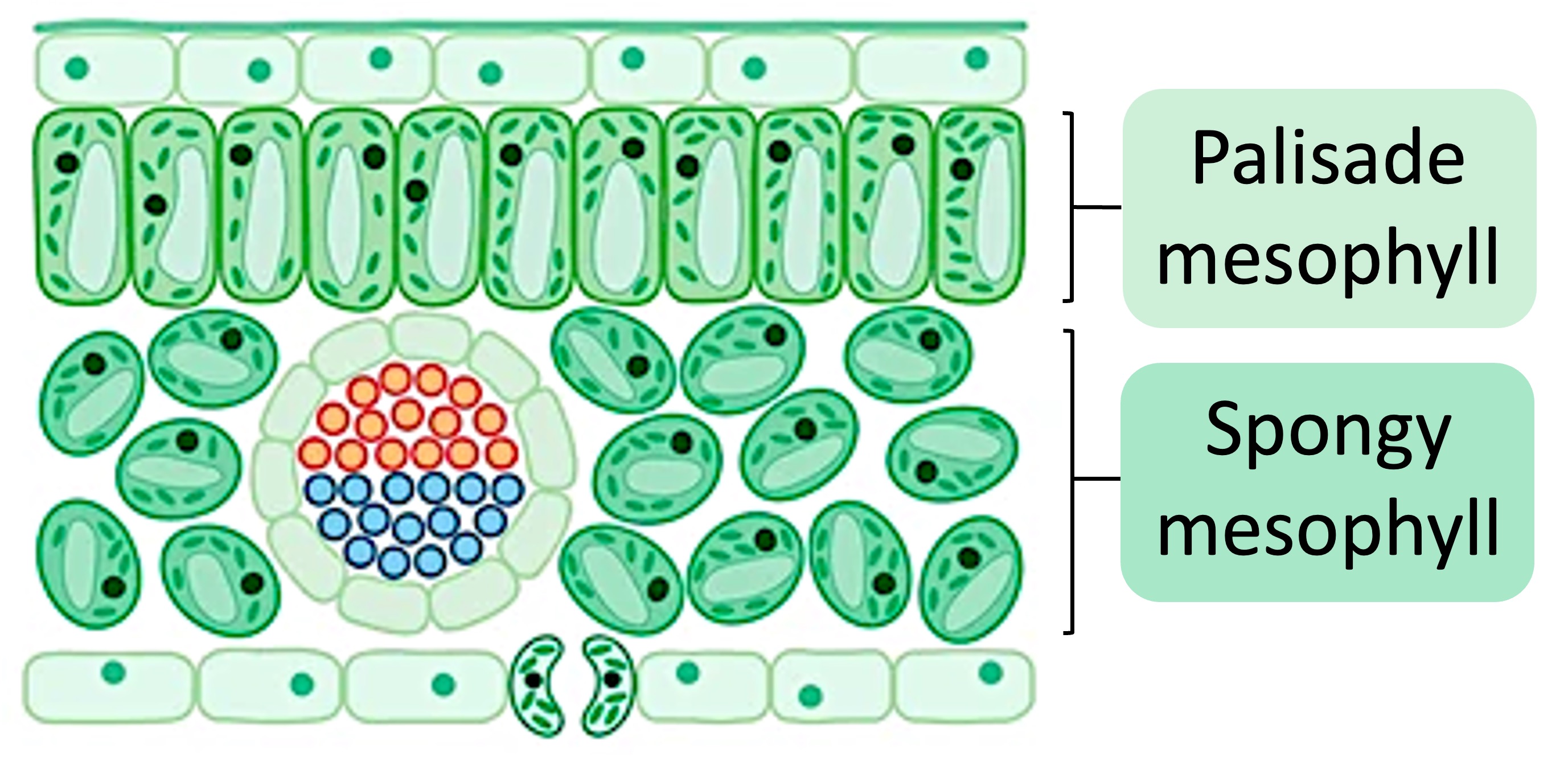
The interior of a leaf is composed of a layer of tissue called the mesophyll – which is organised into two sections:
The cells of the palisade mesophyll are tightly packed and many chloroplasts (for photosynthesis)
The cells of the spongy mesophyll are loosely packed cells that allow circulation of gases
Leaves have vascular tissue such as xylem and phloem
The xylem functions to transport water and minerals from the roots of the plant (via transpiration)
The phloem transports dissolved sugars produced by photosynthesis to other parts of the plant (as sap)
3.1.8 Distribution of tissues in a leaf
The tissues in a leaf are specifically distributed to maximise the processes of photosynthesis and gas exchange
The waxy cuticle covers the exterior surface in order to prevent water loss from the leaf (except via stomata)
The palisade mesophyll is located on the upper half of the leaf (facing sunlight) to maximise light absorption
The spongy mesophyll is located on the lower half (near stomata) and contains air spaces for gas exchange
The stomata are on the underside of the leaf to prevent obstruction and maintain an open channel for gases
The vascular bundle is located centrally to allow for optimal access by all leaf tissue (palisade and spongy)
3.1.9 Transpiration as a consequence of gas exchange in a leaf
Transpiration is the loss of water vapour from plants
Light energy converts the water within the leaves into vapour, which then evaporates and diffuses from the leaf via the stomata
New water is absorbed from the soil by the roots, creating a difference in pressure between the leaves (low) and roots (high)
Water will flow – via the xylem – along the pressure gradient to replace the water lost from the leaves (transpiration stream)
The rate of transpiration by a plant can be affected by a number of factors:
Sunlight – Light stimulates the opening of stomata, as gas exchange is required for photosynthesis to occur
Temperature – High temperatures increase the vaporisation of water, leading to greater diffusion from the leaf
Wind – The flow of air functions to remove the water vapour surrounding the leaf, increasing diffusion rates
Humidity – A higher concentration of water vapour in the air will decrease the rate of diffusion from the leaf
3.1.10 Stomatal density
The amount of water lost from the leaves (transpiration rate) is regulated by the opening and closing of stomata
The opening of the stomata is controlled using guard cells, which are found in pairs on either side of a stoma. By opening and closing the stoma caused by pressure changes the amount of water lost through transpiration can be minimized.
(A closed stoma - guard cell with low pressure in the cytoplasm. An open stoma - guard cell with high pressure in the cytoplasm)
Transpiration rates will be higher when stomatal pores are open than when they are closed (gas exchange impacts transpiration)
Variations in the density of stomata may arise due to genetic factors or growth under different environmental conditions
Higher level (check if i have to know oxygen dissociation curves)
3.1.11 Adaptations of foetal and adult haemoglobin for the transport of oxygen
Haemoglobin function - Haemoglobin is a protein found in red blood cells that binds to oxygen in the lungs and transports it to tissues and organs throughout the body
Haemoglobin structure - Haemoglobin is a conjugated protein. A conjugated protein contains both a polypeptide chain (protein portion) and a prosthetic group (non-protein portion).
Affinity - Affinity refers to the strength of the interaction or binding between two molecules. High affinity = strong attraction, tightly bound together. Low affinity = loosely bound together, break apart easily
A developing fetus needs to get oxygen and nutrients to its tissues, but the physiology of a fetus differs dramatically from an adult. Mostly, it is the lack of functional lungs. This fact makes it necessary for a fetus to have adaptations to its haemoglobin so that more oxygen can be transported from the placenta to the fetus’ circulatory system.
During pregnancy a foetus obtains oxygen via the placenta and the fetal umbilical cord which transports the blood. O2 dissociates from haemoglobin in maternal blood in the placenta and binds to haemoglobin in fetal blood.
Inside the maternal blood, there is generally a higher concentration of o2 and a lower concentration of co2. This creates a pressure gradient that allows o2 to naturally flow into the fetal blood and co2 to flow out of the fetal blood.
Humans produce different types of haemoglobin before and after birth. At birth, a baby still has red blood cells with fetal haemoglobin for several months. Fetal haemoglobin has a stronger affinity for oxygen than adult haemoglobin.
3.1.12 Bohr shift
In very active tissues, for example during physical exercise, co2 from cellular respiration diffuses into the blood stream.
Most of it diffuses into red blood cells where it reacts with water (with the help of an enzyme) forming carbonic acid (h2co3). The acid dissociates into H+ and HCO3- ions and diffuses into the blood plasma.
Carbon dioxide lowers the pH of the blood (by forming carbonic acid), which causes haemoglobin to release its oxygen. This is known as the Bohr effect– a decrease in pH shifts the oxygen dissociation curve to the right.
Due to the increase in co2 release the pH becomes lower. This change in pH causes structural changes in haemoglobin. Hence haemoglobin is promoted to release its oxygen at the regions of greatest need.
3.1.13 Oxygen dissociation curves as a means of representing the affinity of haemoglobin for oxygen at different oxygen concentrations (LOOK AT MORE)
An oxygen dissociation curve is a graph showing the percentage saturation (of oxygen) of haemoglobin against the concentration of available oxygen. The concentration of oxygen is usually shown as partial pressure of oxygen in air using the units Pascal (kPa).