Week 3: MHC and Antigen Presentation
Learning Outcomes:
What sort of antigens do lymphocytes “see”
How do B cells “see” their antigens
How do T cells “see” their antigens
MHC restriction
What are MHC molecules?
general properties
structure
function
Cellular expression
How do antigen presenting cells acquire their antigens and process them to T cells?
The MHC processing pathways
Antigens can 'see’:
B lymphocytes
Whole proteins, carbohydrates, lipids, nucleic acids, peptides
Antigens on cell-membranes or in solution
Can be seen by anything with a B-cell receptor in any situation
T lymphocytes
Small peptides presented in association with Major Histocompatibility Complex (MHC) molecules only.
If peptides are too big they won’t fit in in the MHC
T cells are MHC restricted
Can only see peptide antigens when they are presented on MHC molecules
Peptides are cell-associated – do not see free floating/soluble antigens
T cells are produced in the bone marrow and mature in the thymus
Naive T cells = T cells that have not encountered any specific antigen
Naive T cells then go into the blood stream
Naive T cells recirculate in lymphoid organs to allow them to encounter foreign antigens
Naive T cells for specific antigens are very rare
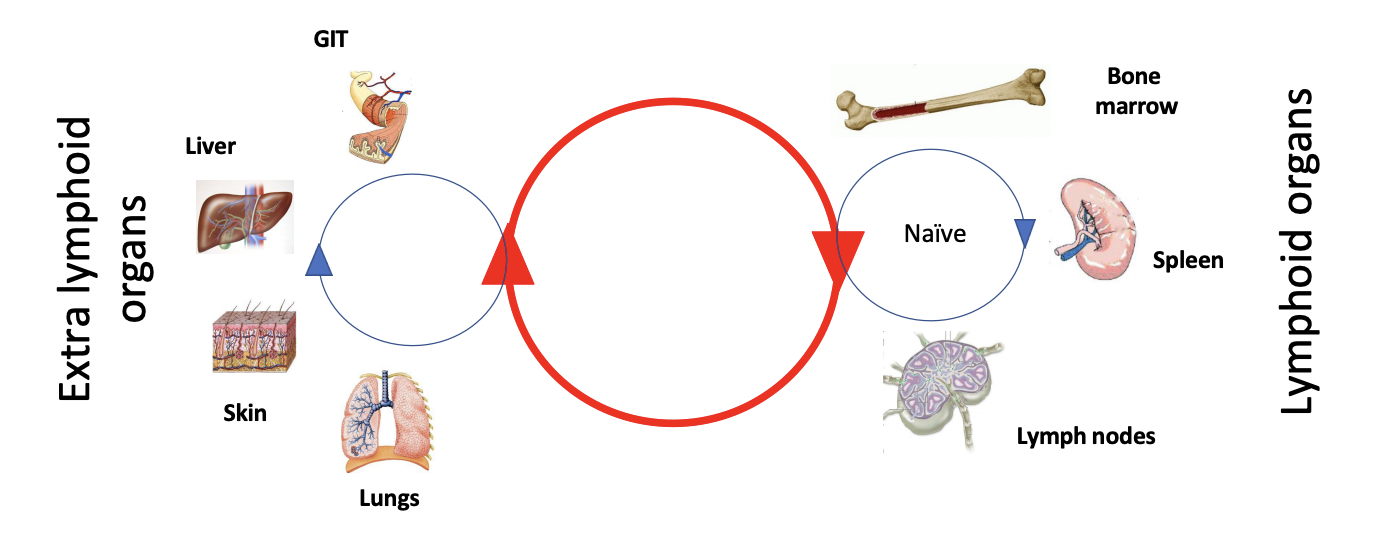
Naive T cells need to meet other cells that will maximise the chance of encountering antigens – e.g. dendritic cells
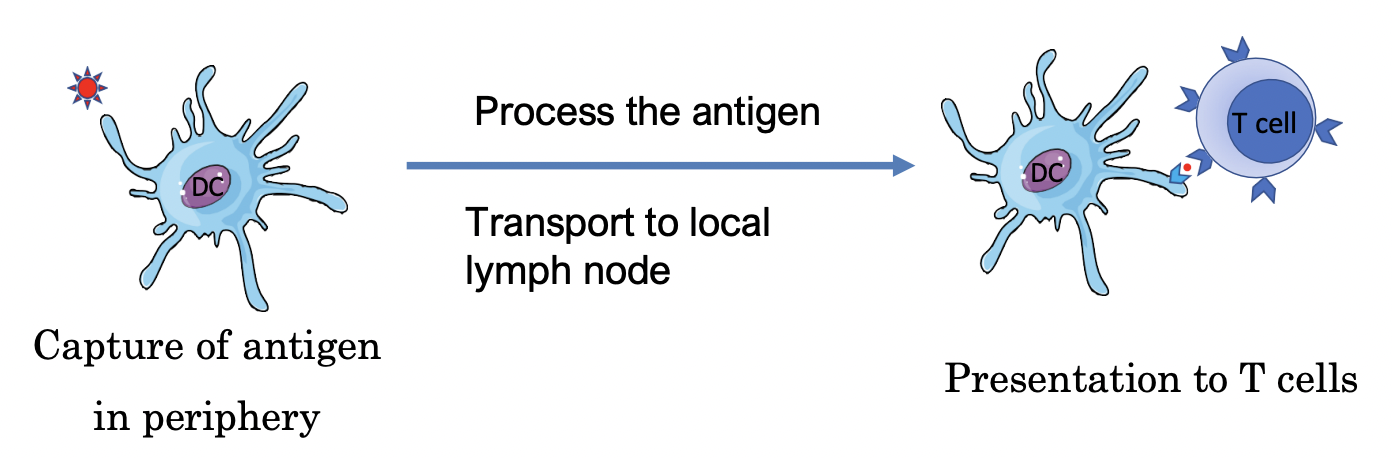
Process of Antigen Presentation to Naive T Cells:
Immature dendritic cells (DC) capture an antigen presenting cell and activate it.
DC travels down afferent lymphatic vessel through lymph until it reaches the lymph nodes, with the DC itself maturing while it is migrating
Immature DC are better at phagocytosis and capturing antigens
Mature DC are better at transporting and presenting antigens
Fully mature DC can present the antigen to T cells
Structure of Lymph Nodes:
Afferent lymphatic vessel is always at the periphery
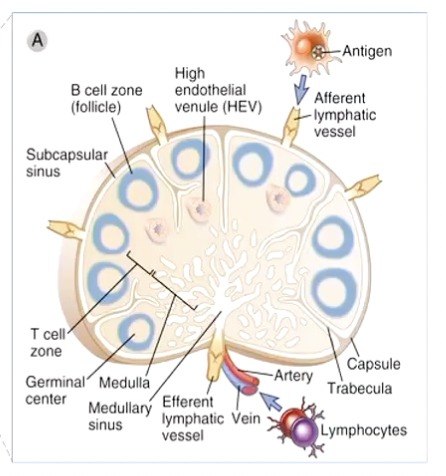
Then goes to the cortex → paracortex
Paracortex is deeper inside the cortex and contains T cells that interact with dendritic cells
Paracortex → medulla
Dendritic cells presenting antigens will be free floating and drained with the movement of lymph
High Endothelial Venule (HEV):
Highly specialised blood vessels that connects the allow lymphocytes in the blood to go directly to the lymph nodes
Endothelial cells that are cuboidal
Circulation of Lymph:
From periphery via afferent vessels
Lymph is filtered
Returns via the efferent vessels
Filtered lymph is returned back to the blood
Dendritic Cells:
Peripheral tissues contain different types of dendritic cells in specific locations with different functions
DC are strategically located to maximise chance of first encounter
DC at peripheral sites express a range of PRRs (pattern recognition receptors)
Allows them to detect microbial patterns or PAMPs (Pathogen Associated Molecular Patterns)
Triggers production if inflammatory cytokines
Triggers uptake of antigens (phagocytosis)
DC are ‘professional antigen presenting cells’:
Only DC can activate naive T cells
Antigen Processing:
After the antigen is picked up
Big captured proteins need to be broken down into peptides by proteases
Smaller peptide antigens are loaded onto special surface molecules so that T cells can see them
These molceules are the MHC molecules
Dendritic Cell Maturation:
As DC migrate from periphery to lymph node, they mature
Characterised by modification of their cell surfaces
Upregulation of many molecules:
MHC
CD86
CD80
Cytokines
Adhesion molecules
Dendritic Cell Migration:
DC in the periphery need to meet with naive T cells which are located in particular areas of the lymph nodes
Antigen-specific receptors on T cells bind only small parts of molecules from pathogens
a few amino acids only
These are called antigenic determinants or epitopes
Small, specific part of the antigen molecule which the T cell/antibody attaches to
T cells recognise processed antigens
Only linear parts are able to be seen by T cells (in blue)
Those peptides, after they are cut out of the original protein, need to fit into the groove of MHC molecules
Too large - won’t fit into groove

This happens everywhere:
at any type of epithelium
If DC with antigen reaches lymph node – the antigen will be collected
If it reaches bloodstream, then spleen – the antigen will become a blood borne infection
Antigen-presenting cells in the spleen capture the blood borne antigens
MHC Molecules:
Major Histocompatibility Complex
First discovered in tissue transplant studies on mice
Individuals identical (inbred and identical) at MHC locus accept grafts from one another
Individuals dissimilar reject grafts from one another
Physiological functions on MHC proteins is to display peptides (only) to antigen-specific T cells
In all mammalian species there are 2 sets – MHC class I and MHC class II
Genetic region primarily responsible for rapid tissue rejection
In humans, it’s called the Human Leukocyte Antigen (HLA)
Responsible for displaying antigens to T cells
Highly polymorphic – have many different structures
> 20000 MHCI
> 7000 MHCII
MHC Genes
Class I and Class II are the most polymorphic genes in any mammalian genome
Polymorphism = variation between individuals
MHC genes are co-dominantly expressed – 1 gene from mother and 1 gene from father and each gene is expressed equally
3 loci with each class I gene: HLA-A, HLA-B, HLA-C
Hence, 3 maternal and 3 paternal HLA molecules will be expressed
6 on every dendritic cell
Genes are located on short arm of chromosome 6
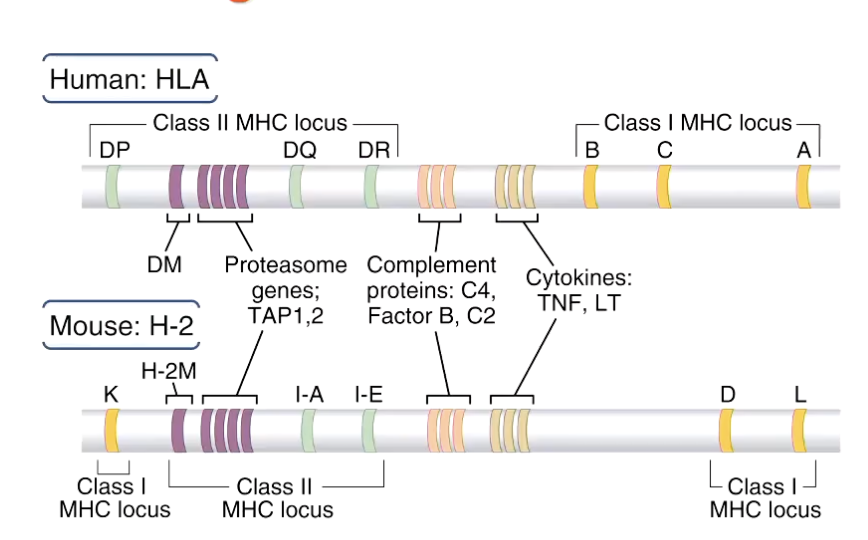
In human system, genes encoding for the proteasome and transport proteins are located close to the MHC molecules
So are cytokines, TNF (tumour necrosis factor) and LT (lymphotoxin)
MHCI are expressed by all nucleated cells
Thats why graft rejection can happen with any tissue with a different recipient
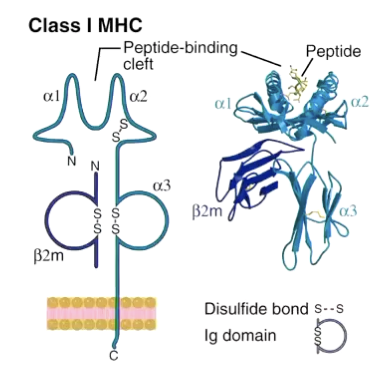
Made of only 1 alpha chain with different domains
Alpha 1 and 2 domains are highly variable
Create a groove/cleft where the peptide can go
Alpha chain is connected to the plasma membrane
Associated with beta-2m (molecule) which is not connected to any signalling structure
Variable top region, stable/constant bottom region
Constant alpha-3 can only pick up CD8
Only can activate CD8 T cells
MHCII are only expressed only on APC (antigen presenting cells), both professional and non-professional
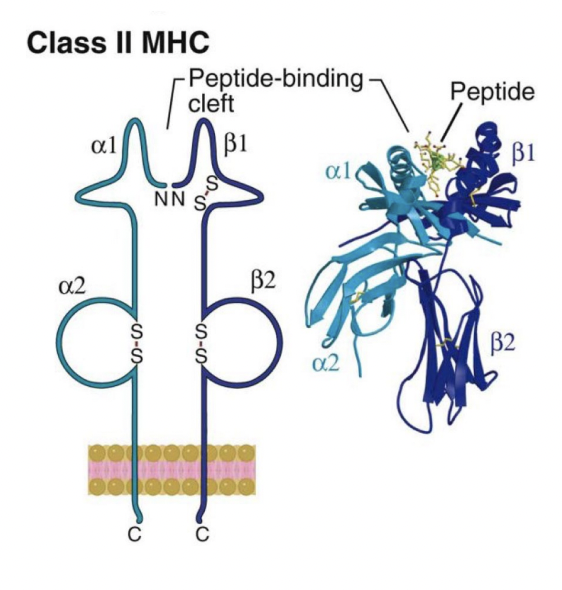
Made of 2 chain – alpha and beta chain
Highly variable region at the top – create a cleft for peptide to go
Constant region (region closest to the plasma membrane) can only stimulate CD4 T cells
Invariant region of both MHC molecules determine what type of T cell is stimulated:
CD8 – MHCI
CD4 – MHCII
General Properties of MHC Molecules:
Highly polymorphic – different individuals are able to present and respond to different microbial peptides
Co-dominantly expressed (both parental alleles of MHC gene are expressed) – increases number of different MHC molecules that can present peptides to T cells
MHC-expressing celltypes:
Class II (APC: DC, macrophages, B cells, endothelial cells) – CD4+ helper, T lymphocytes interact with DC, macrophages and B lymphocytes
Class I (all nucleated cells) – CD8+ CTLs (cytotoxic T lymphocytes) can kill any type of virus infected cell
Also expressed on platelets (non-nucleated) because they come from nucleated megakaryocytes
MHC Peptide Binding Clefts:
There is a precise fitting between a peptide and MHCI and II groove
No more than 10-11 amino acids
MHC Restriction:
CD8+ T cells will only react to peptides on the class I molecule
CD4+ T cells are only able to see peptides on the class II molecule
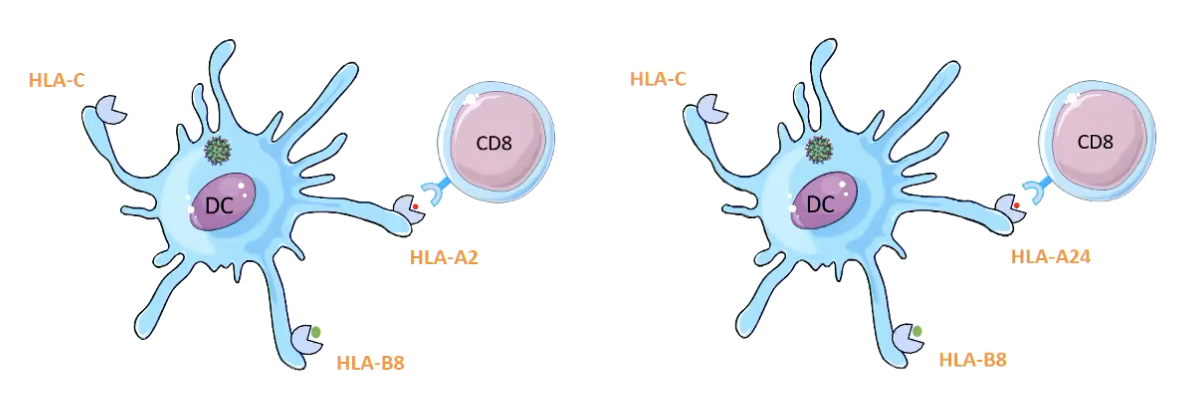
Specificity of HLA molecule itself is also involved:
A HLA-A2 restricted response will only occur when the CD8+ T cell binds to the HLA-A2 molecule
No response if it tried to bind to HLA-A24 molecule
Significance of MHC Diversity:
Have the possibility to present in different way
Proteasome will be chopping big proteins into small peptides
Those peptides will either be presented on class I or class II molecules
MHC polymorphism evolved because it ensures that individuals will be able to deal with diverse microbes – population will be protected by merging infections.
Antigen Processing Pathways:
Extracellular antigens will always go to class II
Intracellular antigens will always got to class I
DCs are capable of presenting MHC class I and class II
MHC Class I Pathways:
Antigen uptake
Inside the cell:
virus
tumour
microbial proteins that have been transported out of phagocytosis
Antigen chopping my proteasome
amplified by inflammatory cytokines (TNF, IL-1)
Small peptides transported using TAP (Transporter Antigen processing)
Go into the endoplasmic reticulum
Class I molecules are being synthesised
MHC class I molecule is loaded with the peptide onto cleft
Goes to then exits the Golgi apparatus
Vesicles are exocytosed and reach the surface
= Peptide expressed on class I MHC molecules coming from inside the cell
Stimulate only CD8+ T cells
Usually become cytotoxic T cells
Results in the killing of cells expressing those antigens
Bare Lymphocyte Syndrome is caused by TAP deficiency:
shows TAP are very important for the quality of an immune response
MHC Class II Pathways:
Antigens from outside the cell
Will be phagocytosed of endocytosed
Endocytosis of extracellular protein – creates phagosome
Phagosome fuse with existing lysosomes to form phagolysosomes
Enzymes anf toxic substances in phagosome kills microbes
Nitric Oxide
Reactive Oxygen Species
Some are preformed
Production may be triggered by PRR binding
Goes to the ER
alpha and beta chains of MHC molecule are being synthesised to become more stable (initially unstable)
The invariant chain with CLIP (Class II Invariant Chain Peptide) occupies the binding cleft of the newly synthesised class II molecules
CLIPS keeps MHC II stable
Blocks other peptides from binding the newly synthesied MHC II molecules
Class II molecules are transported out of the ER via the Golgi in an exocytic vesicle
Exocytic vesicle fuses with the phagolysosome
Brings MHC II molecules and degraded proteins togethers
Peptide is presented in the groove of the class II molecule, at the cell surface
CD4+ T cells are able to bind to the surface of APC
CD4+ helper T cells do not kill like CD8+ T cells, but they help by producing cytokines and chemokines which will further activate the APC
Or they activate B cells
amplify antibody production
amplify other B cell functions
Presentation:
Exogenous Antigen → Class II → CD4+ T cells
Endogenous Antigen → Class I → CD8+ T cells
Cross-Presentation:
Exogenous Antigen → cytosol OR vacuole → class I → CD8+ T cells
Questions for Study:
When antigens enter through epithelial barriers, such as the skin or intestinal mucosa, in what organs are they concentrated? What cell type(s) plays an important role in this process of antigen capture?
What are MHC molecules? What are human MHC molecules called? How were MHC molecules discovered, and what is their function?
What are the differences between the antigens that are displayed by class I and class II MHC molecules?
Describe the sequence of events by which class I and class II MHC molecules acquire antigens for display.
Which subsets of T cells recognize antigens presented by class I and class II MHC molecules? What molecules on T cells contribute to their specificity for either class I or class II MHC–associated peptide antigens?