Chapter 5: Antigen Recognition by T Lymphocytes
Gene Recombination in TCRs and BCRs
Somatic recombination is a genetic process utilized by both T-cells (TCRs) and B-cells (BCRs) to create diverse receptors.
Only TCRs and BCRs utilize some form of recombination specifically applied to their receptor genes.
Activation of lymphocytes leads to clonal expansion where they proliferate, a process termed "proliferation." This occurs even post-maturation.
For T cells, somatic recombination happens in the thymus
Comparison of Functions
While antibodies perform their function by binding directly to various soluble antigens, T-cells require direct cell-to-cell interaction for their activation and subsequent immune commands.
The diversity of TCRs is essential to suit various antigenic challenges; this is achieved through somatic recombination within T-cells, similar to B-cell development but without further RNA changes post-maturation.
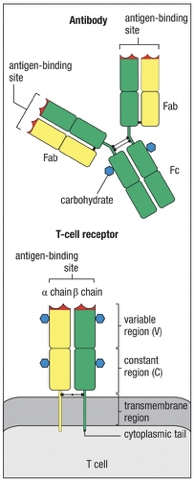
T-cell Receptors (TCRs)
TCRs are never secreted; they remain on the cell surface, mediating cell-to-cell interactions.
TCRs recognize specific peptide-MHC complexes presented by other cells, leading to effector functions predominantly via cytokine secretion (e.g., interferon-gamma from TH1 cells).
While TCRs interact with antigens, their recognition mechanism is fundamentally different from antibodies, necessitating the presence of MHC.
TCRs bind to one type of antigen - protein
Requires presentation by another human cell
Ag-presenting glycoproteins (MHC molecules)
MHC (Major Histocompatibility Complex)
Definition: MHC molecules are membrane-bound glycoproteins that present peptide fragments to T-cells.
MHC Class I: Expressed on all nucleated cells and important for presenting internal antigens to CD8+ cytotoxic T-cells.
MHC Class II: Found only on antigen-presenting cells (APCs) like dendritic cells, playing a crucial role in presenting external. antigens to CD4+ helper T-cells.
The compatibility of MHC molecules is essential in transplantation; mismatched MHC can lead to graft rejection due to recognition of foreign tissue by the immune system.
Large number genetically determined variants (alleles) in human population. Polymorphism
i.e. differences between individuals in the MHC molecules.
Transplant Implications
MHC compatibility is a major determinant in successful organ transplants. The presence of different MHC alleles among individuals necessitates careful matching to prevent rejection.
Matching Process: Advances in technology have facilitated the matching process for organ transplants, improving patient outcomes considerably
TCR Structure/Mechanics
Membrane bound glycoprotein
Resembles a single antigen-binding arm of immunoglobulin.
Consists of two polypeptides the and chains; one Ag-binding site.
“membrane bound”; no secreted form
No differing effector functions
Each chain has variable region (binds Ag) and a constant region just like Igs.
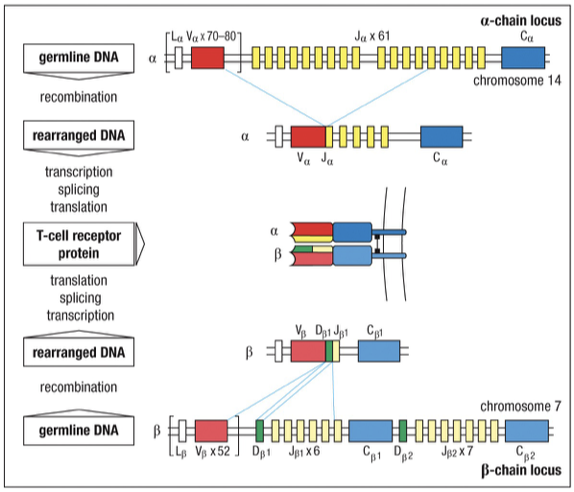
The alph chain contains V and J segments (like Ig Light-chain)
The chain contains V, J, and D segments (like Ig Heavy-chain)
TCR Diversity/Development
Gene rearrangement = sequence variability in V regions (similar to B-cell)
No further mutation in Ag-binding site after Ag stimulation.
No switching of constant-region isotype.
TCRs exhibit a high diversity conferred through genetic recombination that creates a nearly infinite number of receptor specificities.
Mechanisms: TCR diversity arises from V(D)J recombination, similarly to BCRs, involving complex interactions among gene segments (V, D, and J segments).
Rearrangements occur during T cell development in thymus; mature T cell has one functional chain, one functional chain, together they define a unique TCR.
An individual's population of T-cells = many millions of different TCRs, each defines a clone of T-cells with a single Ag-binding specificity.
Effector functions handled by T-cell itself
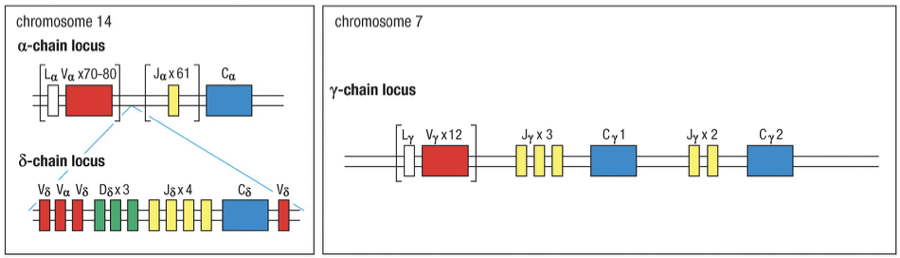
Human TCR alp chain locus (Chr 14)
Human TCR -chain locus (Chr 7)
chain contains two D sections, therefore TCR β chains have a finite maximum (~4 attempts, 2 per maternal and paternal chromosome)
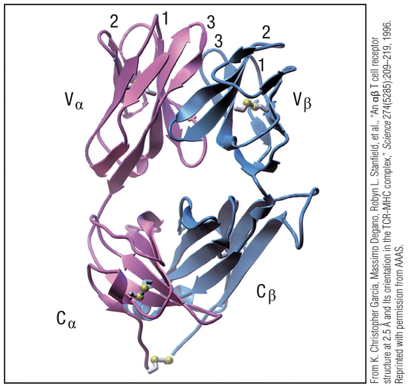
HVs/CDRs (sequence variation) of TCR
Sequence variation in the and chains clustered into regions 3 of “hypervariability” region.
Correspond to beta pleated sheet loops of the polypeptide farthest from the T-cell membrane.
Loops = Ag-binding site; complementarity-determining regions (CDR1,CDR2,CDR3)
TCR Binding
TCRs possess single binding site for antigen.
Used only as “cell-surface” receptors for Ag.
Never as soluble Ag-binding molecules
Occurs in context of two opposing cell surfaces.
Multiple copies of TCRs bind to multiple copies of Ag:MHC complex on the opposing cell.
Multipoint attachment
TCR Complex
The TCR complex is composed of 8 polypeptides.
and chains form core.
the constant chains interact with intracellular signaling molecules
Expression of the TCR on the cell surface requires association with additional proteins
TCR = : heterodimer
Alone can not leave Endoplasmic reticulum
: heterodimer stable association with 4 “invariant” membrane proteins
CD3 complex (Chromosome 11)
homologous CD3
CD3
CD3
also contain
chain (Chromosome 1)
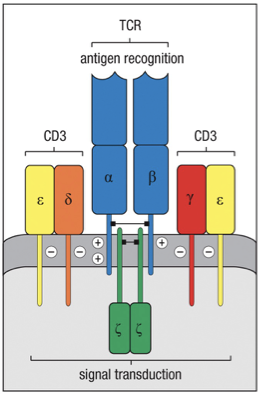
TCR has short cytoplasmic tails.
= gamma
= delta
= epsilon
= zeta
TCR has short cytoplasmic tails.
Therefore, CD3 complex & chain, longer sequence, transduce signals to cell’s interior after Ag recognition by TCR.
Lack of functional CD3 & CD3 = low TCR expression and impaired signal transduction = immunodeficiency.
Gamma: Delta T Cell Class
: TCR structure similar to : TCR
chain resembles chain
chain resemble chain
A cell can express either or TCRs, never both.
Cells expressing : TCR are called : T cells and cells expressing : TCRs are called : T cells.
Cells expressing : receptors form a small subset of all T cells (only 1-5%).
Can be dominant T cell population in epithelial tissue.
: T-cells immune functions and Ags less well defined.
Generally not associated with recognition of antigen through MHC.
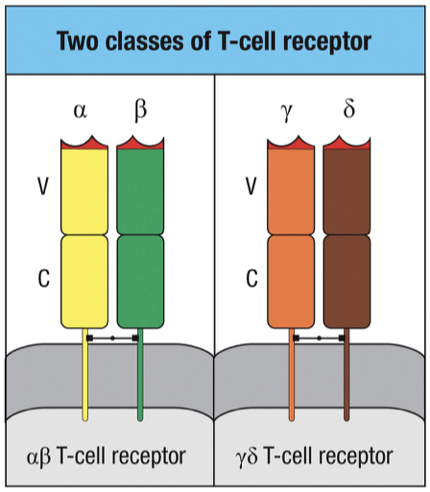
TCR Classes
Alpha-Beta T Cell Receptors (TCRs)
Composed of alpha and beta chains.
Majority population of T cells (95-99%).
Function: adaptive immune response.
Requires co-receptor (CD4/CD8) and MHC presentation for antigen recognition.
Gamma-Delta T Cell Receptors (TCRs)
Composed of gamma and delta chains.
Minority population of T cells (1-5%).
Function: innate immune response.
Do not require MHC for antigen recognition and do not form memory cells.
Chromosoal Organization
Chromosomal Location
Alpha Chain Locus: Located on chromosome 14.
Delta Chain Locus: Entire gene locus is embedded within the V and J sections alpha locus on chromosome 14.
Rearrangement of V and J segments in alpha chain results in excision of delta locus, disabling gamma-delta TCR formation from that gene.
Rearrangement and Junctional Diversity
Delta chains can increase diversity due to potential inclusion of additional D segments, enabling V-D-D-J configurations.
B Cells vs T Cells (analytic analogy)
B cells = Dogs 🐶 → sniff things directly
Dogs can walk around and sniff anything they find.
They do NOT need someone to present the smell to them.
Dogs sniff freely → B cells recognize antigen freely (NO MHC needed), use epitope and paritope relationship
T cells = Cats 🐱 → must be shown the antigen
Cats don’t randomly sniff everything. Instead, imagine cats only react when:
Someone brings them an object on a tray and shows it to them.
Cats must be shown → T cells require antigen presentation on MHC
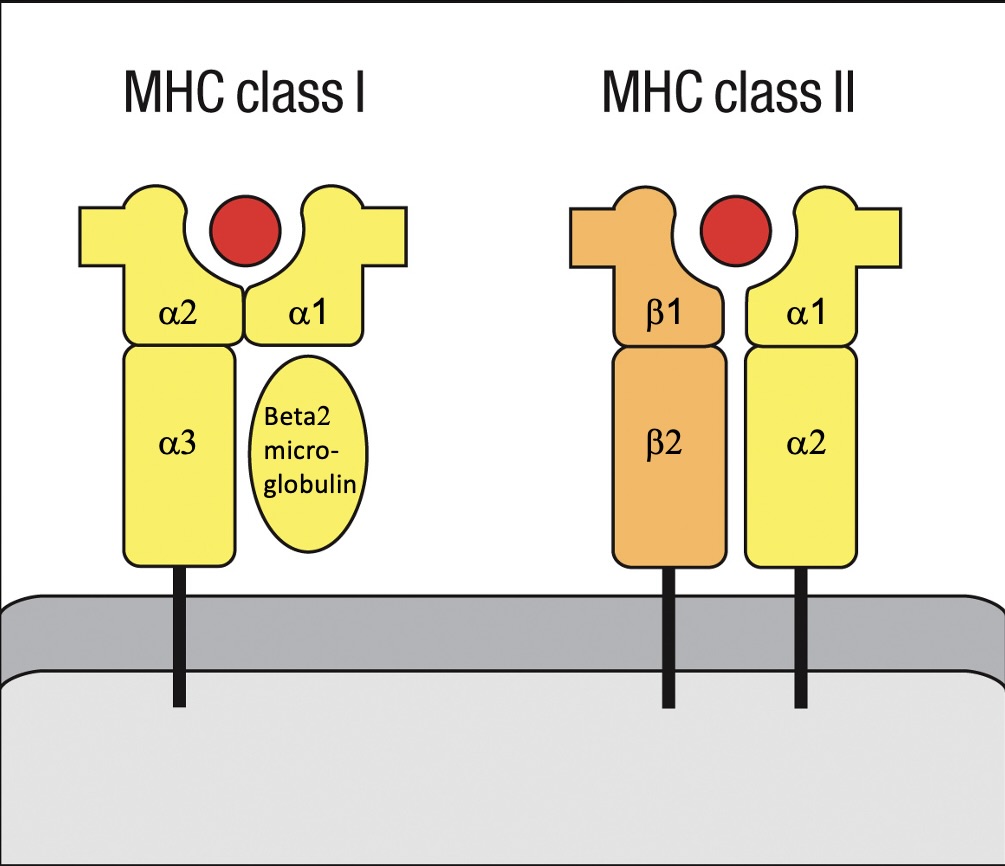
Structure of MHC Molecules
MHC Class I
Presents intracellularly processed peptides
Engages with CD8 T cells.
Structure includes an alpha chain that forms a groove for peptide binding and a circular beta-2 microglobulin.
MHC Class II
Presents extracellularly derived peptides
Engages with CD4 T cells.
Structure features alpha chain and beta chain (2 alpha and 2 beta subunits), with longer binding grooves allowing peptide extensions beyond the ends of the pocket.
Peptide Binding of MHC
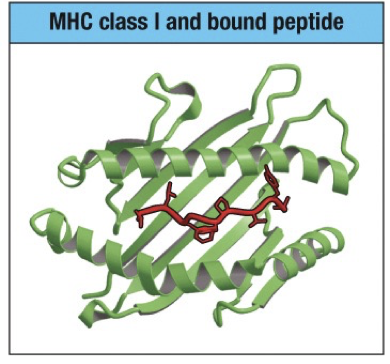
MHC Class I
Length limited because the two ends of the peptide are grasped by pockets situated at the ends of the peptide-binding groove.
Binding groove is composed of alpha helices above a beta-pleated sheet.
8, 9 or 10 amino acids
Restriction: Peptides are bound with hydrophobic or basic residue at the carboxyl terminus (protein runs from N to C terminus).
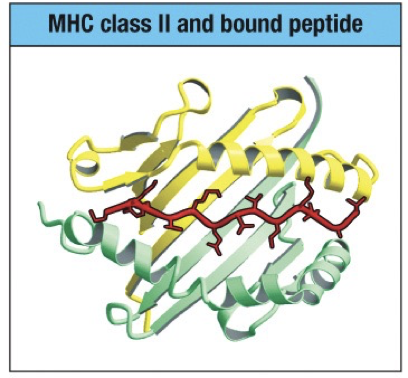
MHC Class II
Two ends of the peptide not pinned down into pockets at each end
Extend out at each end of the groove
Longer and more variable in length than peptides bound by MHC class I
10-25 amino acids in length or longer
No binding restriction
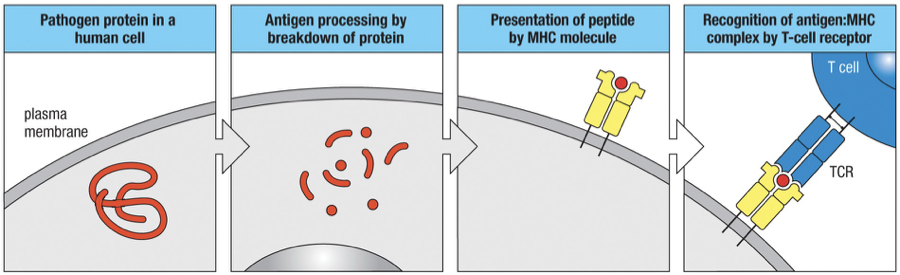
Generic MHC Processing
Ag Processing: Pathogen-derived proteins must first be broken down into peptide fragments.
Ag Presentation: Peptides are displayed on cell surfaces bound to MHC. For MHC Class I and II, the differences lie in their processing pathways and peptide lengths.
MHC I required specific peptide binding at defined lengths; MHC II allows extensions of peptides beyond the binding groove.
MHC I Pathway (Endogenous Pathway)
What MHC I presents:
Peptides from proteins made inside the cell
Example: viral proteins, tumor proteins
Who can use MHC I:
Any nucleated cell
Steps:
Protein degradation in cytosol
Proteins are cut into peptides by the proteasome (including β1i, β5i).
Transport into ER
Peptides enter the endoplasmic reticulum (ER) through a transporter called TAP.
Binding to MHC I
MHC I molecules are made in the ER.
Peptides bind to MHC I inside the ER.
Transport to cell surface
MHC I + peptide complex is packaged into vesicles.
Vesicles move to the plasma membrane.
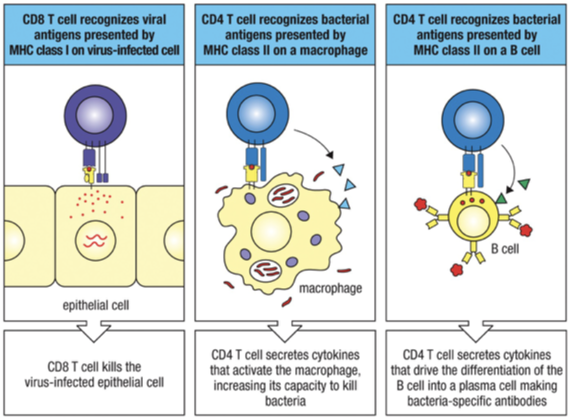
Presentation to CD8 T cells
MHC I presents peptide on surface.
CD8 cytotoxic T cells recognize and kill infected cells.
Simple flow:
Cytosol → ER → MHC I binding → Vesicle → Cell surface
CD4 vs CD8 T Cells
CD4: Defined by CD4 glycoprotein expression
CD8: Defined by CD8 glycoprotein expression
Different functions
Different types of target pathogens
CD8 T-cells are cytotoxic and kill cells that are infected with a virus or other intracellular pathogen.
Prevents pathogen replication and further infection of healthy cells.
CD4 T-cells - help other immune cells respond to extracellular sources of infection.
CD4 T Cell Subclasses
Subclasses of T Helper Cells
TH1 Cells: Activate macrophages, involved in cell-mediated immunity.
TH2 Cells: Stimulate B cells to produce antibodies (e.g., IgE).
Cytokines: Released by T helper cells to mediate immune responses and activate other immune cells.
Proteins Involved in MHC I Pathway
Immunoproteasome Components: β1i and β5i help produce peptides that fit into MHC I (8-10 AA) in the cytosol
Beta 1i = coded LMP2 gene
Beta 5i = coded LMP7 gene
TAP1 and TAP2: Membrane proteins responsible for transporting peptides into the endoplasmic reticulum (ER) from cytosol.
Chaperone Proteins:
Calnexin: First chaperone that stabilizes nascent chains;
Calreticulin and Tapasin: Assist in loading peptides onto MHC Class I molecules before they reach the cell surface.
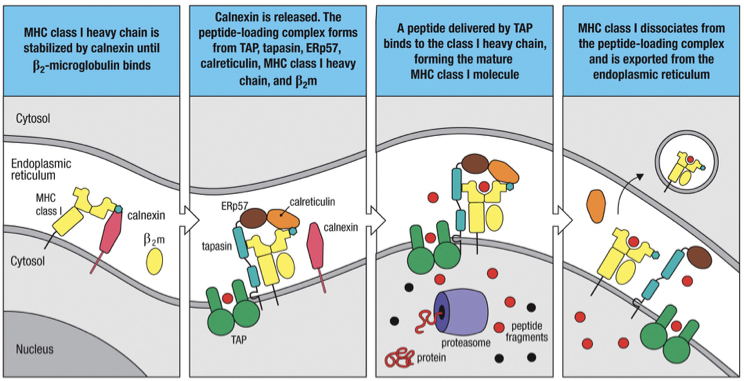
Proteasome (β1i, β5i) → cut peptides
TAP1/2 → move peptides into ER
Calnexin → stabilize heavy chain first (no β2-microglobulin)
β2-microglobulin binds → Calnexin leaves
Calreticulin + Tapasin bind
Calreticulin: stabilizes assembled MHC I
Tapasin: connects MHC I to TAP and facilitates he transport of peptides into the endoplasmic reticulum for loading onto MHC I molecules.
Peptide binds MHC I
MHC I leaves ER → surface
TAP Functionality
TAP1 and TAP2 are integral for peptide transport into the endoplasmic reticulum (ER).
Function: These proteins transport peptides generated by the proteasome to the ER where they load onto MHC Class I molecules.
If either TAP1 or TAP2 is nonfunctional, it can lead to significant immune issues.
Result: Barren lymphocyte syndrome, characterized by a lack of MHC Class I on cell surfaces.
Implication: The inability to effectively present antigens on MHC Class I can hinder the immune system's ability to fight viral and intracellular pathogens
MHC II for APCs (Antigen Presenting Cells)
Unlike MHC I for all nucleated cells to call on CD8 (Killer) T cells, MHC II is specifically expressed by professional antigen-presenting cells, enabling them to interact and activate CD4 T cells through direct recognition of the peptide-MHC complexes.
Professional APCs include:
Dendritic Cells
Macrophages
B Cells
Feedback Loops
APC | CD4⁺ T cell cytokines | T cell subset (if known) | Effect on APC |
|---|---|---|---|
Dendritic cell | IL-2, TNF-α | activated CD4⁺ T cells | Enhances DC survival, maturation, and ability to activate more T cells |
Macrophage | IFN-γ | Th1 | Increases microbial killing, lysosome activity, and cytokine production |
B cell | IL-4 | Th2, Tfh | Stimulates proliferation, differentiation into plasma cells, and antibody secretion |
Dendritic cells migrate to secondary lymphoid tissues (e.g., lymph nodes) after recognizing pathogen-associated molecular patterns (PAMPs) with pattern recognition receptors (PRRs) to activate CD4 T cells (helper cells).
MHC II Pathway (Exogenous Pathway)
Extracellular bacteria, extracellular virus particles and soluble proteins are processed by a different intracellular pathway than intracellular bacteria (cytosolic proteins) and viral proteins.
Vesicles travel inwards from the plasma membrane, their interiors become acidified by the action of proton pumps in the vesicle membrane. Vesicle membranes fuse with other vesicles (lysosomes) to form phagolysosomes, which have enzymes that degrade proteins into peptides.
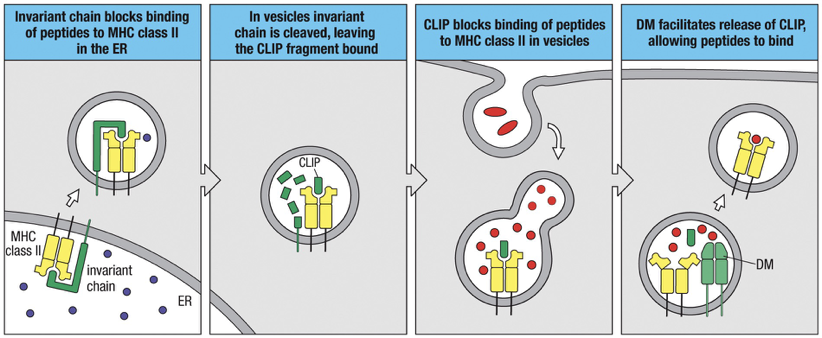
and chains (vary from person) associate in ER with an “invariant” chain (identical in all persons), also called Li
Invariant chain tethers MHC II to ER membrane, blocks peptide binding site (formed by and chain) from binding internal peptides present in the ER, and delivers class II molecules to endocytic vesicles called MHC class II compartments (MIIC).
MIIC is a vesicle that contains proteases (cathepsin S) that selectively cleave invariant chain leaving a small part of the invariant chain to cover the MHC class II peptide-binding groove, also known as Class II-associated invariant-chain peptide (CLIP fragment)
MIIC containing MHC II stabilized with CLIP fuses with phagolysosome (that has the broken down peptides of extracellular material)
HLA-DM inside the MIIC catalyzes the release of CLIP and allows MHC class II molecule to be able to sample peptides
MHC class II molecule samples other peptides until it finds one that binds strongly, binds to appropriate peptide and is transported to cell surface by outward going vesicles.
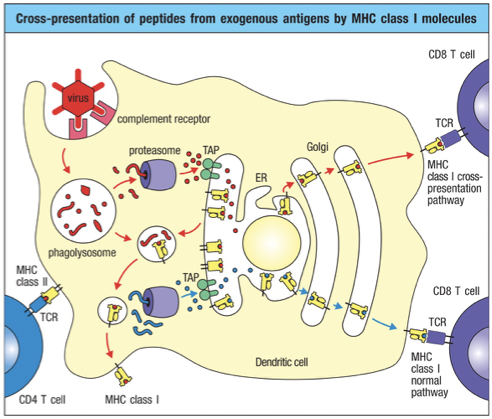
Cross Presentation
Mechanism allowing dendritic cells to present extracellular antigens on MHC Class I molecules.
Underlying need: If a virus avoids infecting dendritic cells, it can evade CD8 T cell recognition.
Cross presentation ensures that even extracellular antigens can lead to CD8 T cell activation when necessary.
The extracellular pathogen peptides just get dumped into cytoplasm and MHC I activation pathway occurs
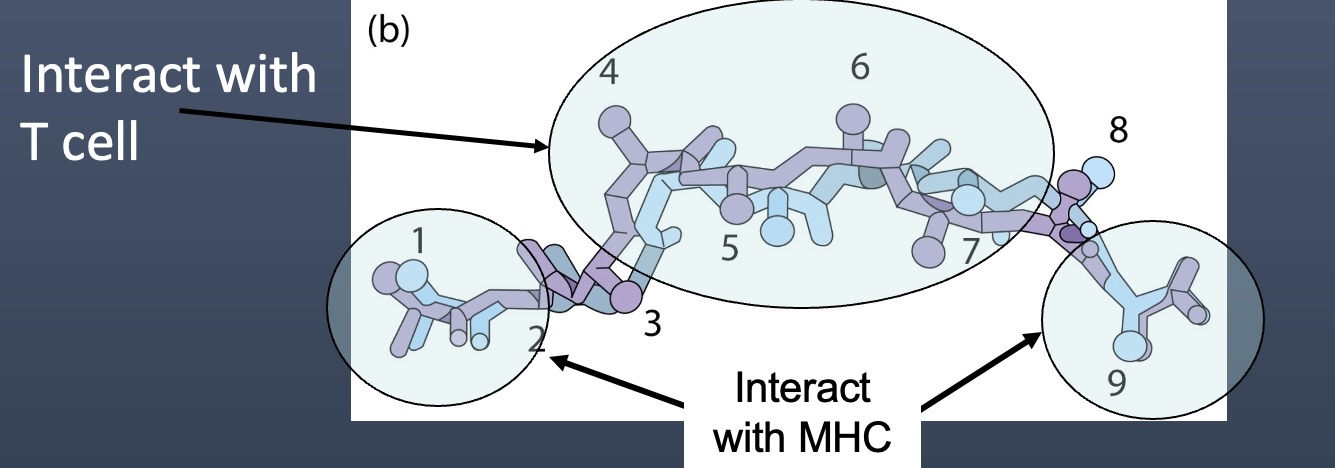
TCR Peptide/MHC Specificity
T Cell Receptors (TCRs) specifically recognize antigens presented by Major Histocompatibility Complex (MHC) molecules.
Binding Mechanism:
The TCR consists of variable regions responsible for antigen specificity, including complementary determining regions (CDRs).
CDR1 and CDR2 bind to Helices of MHC
These regions primarily interact with the MHC molecule. They help determine the specificity of the TCR to the particular MHC isoform.
CDR1 and CDR2 contact the MHC surface itself, ensuring that the TCR can correctly identify and bind to the MHC, which is crucial for stable interaction and T cell activation.
CDR3 binds to Peptide
This is the most variable region of the TCR and plays a critical role in binding to the peptide that is presented by the MHC.
The peptide-MHC complex is highly specific, with the CDR3 providing the necessary flexibility and diversity to accurately bind to a wide range of peptide sequences.
This tripartite interaction (TCR - CDR1/CDR2 with MHC and CDR3 with peptide) is essential for T cell activation and subsequent immune response.
Ends of peptide anchor into MHC pockets
These are called anchor residues and interact with the MHC
The exposed middle of the peptide faces upward
Interacts with TCR
Major Histocompatibility Complex (MHC)
MHC is crucial for determining tissue compatibility and immune response efficacy in transplantation.
Discovery linked back to immunological challenges faced in organ transplant and skin graft issues seen in war pilots.
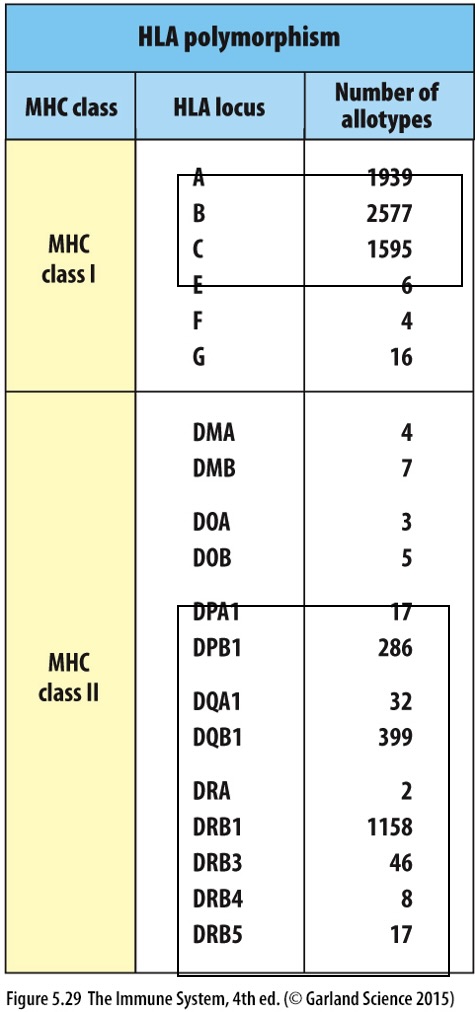
MHC polymorphism contributes to the diverse antigen presenting capabilities among individuals.
Human Leukocyte Antigen (HLA)
HLA = Humen Leukocyte Antigen = MHC for humans
Human Leukocyte Antigen (HLA): Comprises a set of genes encoding for MHC/HLA proteins involved in the immune system's ability to recognize foreign molecules. HLA genes are located on chromosome 6.
These are the HLAs involved in peptide presentation to T Cells
HLA Class I Genes:
Include HLA-A, HLA-B, HLA-C
These genes encode for MHC Class I molecules that present endogenous (intracellular) antigens to CD8 T-cells.
HLA Class II Genes:
Include HLA-DP, HLA-DQ, HLA-DR
Encode for MHC Class II molecules that present exogenous (extracellular) antigens to CD4 T-cells.
HLA Diverisity
Heterozygosity: Refers to having two different alleles (due to both paternal and maternal) at a particular gene locus.
Maximum concentrations of HLA alleles include (for heterozygous):
MHC/HLAClass I (6 alleles):
2 HLA-A alleles
2 HLA-B alleles
2 HLA-C alleles
These code for alpha chain (a1,a2,a3) only of MHC I (beta-2 microglobulin is separate)

HLA Class II (12 alleles):
Because these code for
Each HLA-DP consists of an alpha and a beta chain (2 alleles each).
Each HLA-DQ consists of an alpha and a beta chain (2 alleles each).
Each HLA-DR consists of an alpha and a beta chain (2 alleles each).
These code for alpha chain (a1,a2) and beta chain (b1,b2) of MHC II

Thus, for HLA Class II:
2 (HLA-DP, alpha) + 2 (HLA-DP, beta) + 2 (HLA-DQ, alpha) + 2 (HLA-DQ, beta) + 2 (HLA-DR, alpha) + 2 (HLA-DR, beta) = 12 alleles.
In total, this means a heterozygous person can have:
6 different HLA Class I alleles
12 different HLA Class II alleles.
Additional DRB chains can also pair with DRA
Each HLA gene locus has different allotypes or versions leading to polymorphism
Understanding HLA/MHC Molecules
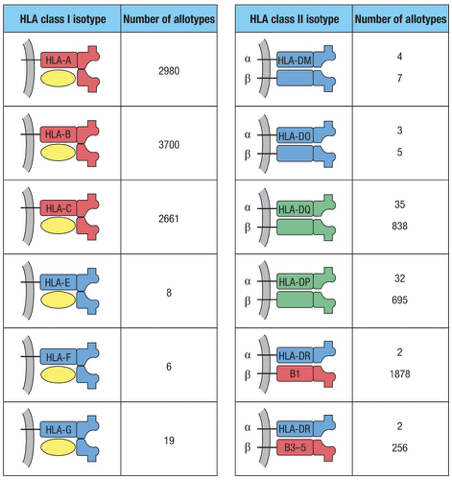
Class I
Presenting Molecules: HLA A, B, C
Actual presenting molecules for immune response.
Involved in presenting peptides to T cells.
HLA E, G, F
Not a presenting molecule for antigen presentation.
Serves as an inhibitory ligand for Natural Killer (NK) cells.
Expression: All nucleated cells that produce MHC class I (HLA A, B, C) will express HLA E on their surface.
HLA-E, HLA-G, HLA-F are also oligomorphic and form inhibitory ligands for NK-Cell receptors
Mechanism: When a cell presents MHC class I molecules, a part of the leader peptide is incorporated into HLA E.
Importance: Infected cells may downregulate MHC class I presentation to evade immune detection; hence, if MHC class I levels drop, HLA E also decreases, making the cell more susceptible to NK cell attack.
Class II
Presenting Molecules: HLA DP, DQ, and DR are examples of MHC Class II molecules responsible for presenting antigens to CD4 T cells.
Functionality: These molecules are involved in both the unloading and loading of peptides, enabling efficient antigen presentation.
HLA-DM and HLA-DO are intracellular and regulate peptide loading of HLA-DP, HLA-DQ and HLA-DR.
Gene Cluster of MHC
Genes that encode MHC class I heavy chain and MHC class II alpha and beta chains are closely linked on a region called MHC Complex (on chromosome 6)
Region called the MHC complex because first identified as region containing polymorphisms underlying graft rejection.
2-microglobulin and invariant chain are not polymorphic (located on chromosomes 15 and 5, respectively).
Key Structures:
HLA class I typically includes a heavy chain (3 sectioned alpha chain shaped like upside down L), and beta-2 microglobulin
HLA class II includes alpha and beta chain with an Invariant chain associated
Polymorphism
Definition: Refers to the presence of multiple allele variants of the same gene across a population, leading to a wide array of protein variants.
Class II diversity contributed by and chains polymorphism
Class I diversity contributed by the one heavy upside down L shaped () chain polymorphism
The DR Gene
Human MHCs differ in the number of DR genes.
The MHC on every human chromosome 6 carries one gene (DRA) for the HLA class II DR chain and one gene DRB1 for DR
In addition, some MHCs carry other DRB3, DRB4 or DRB5.
Any DR chain can pair with the DR chain to form a class II molecule.
Also any DP chain can pair with the DP chain to form a class II molecule.
Also any DQ chain can pair with the DQ chain to form a class II molecule.

Genetic Organization on Chromosome 6
The MHC class I and class II genes occupy different regions of the MHC.
The positions within the HLA complex of the HLA class I and II genes.
The class I genes for presentation are all contained in the class I region.
The class II genes for presentation are all contained in the class II region.
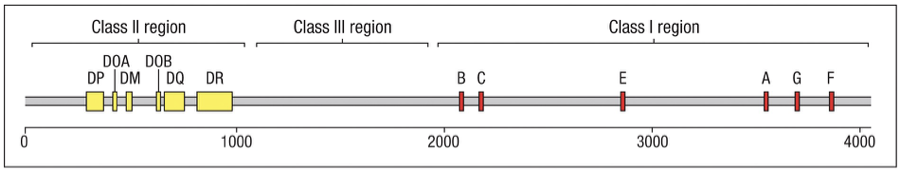
Class III region (central MHC) contains a variety of genes (not shown) involved in other immune functions.
HLA-DP, HLA-DQ and HLA-DR (the and chain genes) are close together and shown as a single yellow block.
HLA-DO ( and genes; DOA and DOB) are separated by the HLA-DM ( and genes together) genes.
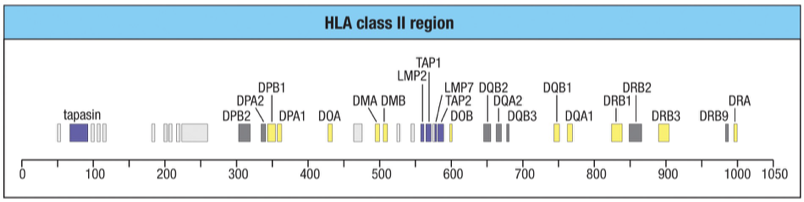
Notice no gene for the invariant chain gene (its on chromosome 5).
MHC I molecules in Class II coding regoion
The class II region includes genes for the peptide transporter (TAP), immunoproteasome components (LMP) and tapasin.
LMP2 = b1i and LMP7 = b5i; subunits in the Immunoproteasome used in MHC I processing (breaking down pathogen peptides)
Variation in HLA allotypes
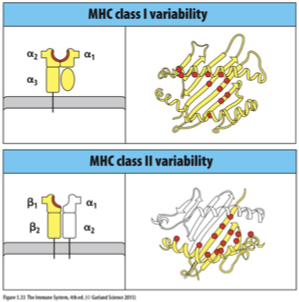
HLA class I molecule allotype variability is clustered in specific sites within the a1 and b2 domains.
These sites line the peptide-binding groove, lying either in the floor of the groove where they influence peptide binding or in the alpha helices that form the walls, which are involved in binding the TCR.
floor binding groove = peptide binding
wall alpha helices = TCR binding
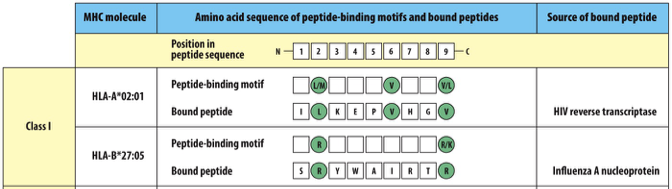
Isoform and Peptide Binding Motifs
Isoform: A different genetic version (allele) of the same HLA gene
Example: HLA-A*02:01
HLA has a molecular clamp with specific “pockets” or anchor residues inside it.
The peptide must have specific amino acids at certain positions to fit.
Those required amino acids = the binding motif
HLA isoforms are genetically fixed.
Pathogens randomly produce peptides.
If a pathogen peptide happens to fit your HLA motif → it gets presented.
If it doesn’t fit → it won’t be presented.
Class I
HLA-A and HLA-B Isoforms: anchor residues at position 2 and 9 (also known as C-terminal)
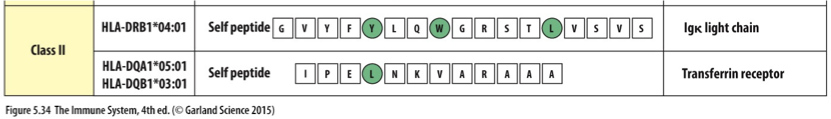
Class II
HLA-DR and HLA-DQ Isoforms are not readily defined
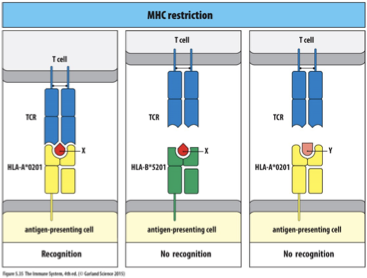
MHC Restriction
MHC Restriction: Concept where TCRs recognize a specific peptide only in the context of a specific MHC molecule.
Experimental Design: Researchers tested TCRs' recognition capability by changing either MHC or peptide while keeping the TCR constant, observing recognition patterns.
If the MHC is changed, the TCR may not recognize the new combination
If the peptide is changed, the TCR may not recognize the new combination
Heterozygous vs Homozygous HLA
Haplotype: With respect of a linked cluster of polymorphic genes, the “set of alleles” carried on a single chromosome.
Every person inherits 2 haplotypes, one from each parent.
Heterozygosity vs. Homozygosity in Genomic Context
Inheritance Patterns: Each individual inherits one allele from each parent, resulting in heterozygosity in most cases unless inbreeding occurs.
Clinical Implications of Heterozygosity: Individuals with greater heterozygosity show stronger immune responses and a more diverse array of presented peptides compared to homozygous counterparts, highlighting the evolutionary advantage of genetic diversity.
Graft Rejection Mechanisms
Host vs. Graft: When the recipient's immune system attacks transplanted tissue due to differing HLAs.
Graft vs. Host: Occurs when the transplanted immune cells attack the recipient's tissues, particularly significant in bone marrow transplants due to the transfer of leukocytes.
Diversity and Disease Resistance
Heterozygous individuals tend to have longer times before developing diseases such as AIDS after exposure to pathogens (like HIV), due to a broader range of MHCs that present varying peptides effectively.
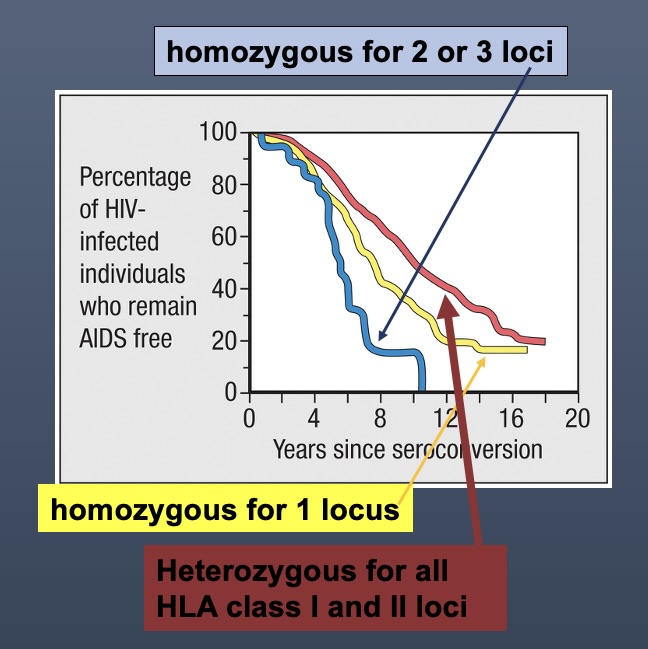
AIDS Diagnostic Criteria
Defined as having fewer than 200 CD4 T cells per microliter, marking a significant downturn in immune function.
The progression from HIV to AIDS varies greatly based on genetic makeup, particularly the presence of heterozygous alleles.