BioChem Test 3 Material
Chapter 14 Digestion
Digestion dosnt generate energy from food, but it consumes the energy. It mainly deals with turning huge ammounts of energy sources and turning it into a usuable forms
For example, proteins are turned into amino acids, complex carbs are turned into monosaccharides
Digestion also abosrobs biomolecules and catalysts
Examples are, Inorganic elements like Sodium, Pottasium, It also abosrbs organic co-factrors ones, vitmain. The absorbtion of inorganic co factors such as iron, copper, ETC
Proteins digestion
Protein digestion produces 4kcal/g
The digestion of prtoein is mediated by a series of molecules called proteases and peptidases, These enyzmes break the peptide bonds which allows the protein to be chopped up and processed easily
The start of the digestions of protein starts within the stomach where the pH of it is very low (very acidic)
How does the low pH of the stomach help with protein digestion?
The low pH of the stomach allows a enzyme called pepsin to be active, since pepsin is activated at very low pH’s. This enzyme helps chop up the proteins in the food in to smaller pieces
The lower pH of the stomach also helps denature most proteins, which makes the proteins unfold and become a more liner shape, which helps the enzymes be able to cut
How this works is the high proton ratio disrupts the ionic bonds between negativley charged and the / groups. The high concentration of Positive H+ protons also disrupts the hydrogen bonds formed between the amino acids residues with other functional groups present
The organs that help in protein digestions are the stomach, Small intesnine, and the pancreas
The digestion through these organs is mediated by hormones
Proteins that are partially digested stimulate the release of a molecule called Cholescystokinin (CCK) and secretin from the small instestin cells
CCK is a hormone which signal that make the pancrease release more protease, such as trypsin, to help complete the protein digestion
CCK also stimulates the gullblader to release bile into the duodenum which will help with the digestion of lipids
Secretin is a horomone released with the stimulas of partially digested proteins, this horomone tells the pancrease to release NaHCO3 to neutrilze the low PH from the digestion products from the stomach, this way it dosnt damage strucutres along its path to complete digestion
Trypsin is an essential enyme in the duodenum
its created by the pancrease in the form of Trypsinogen (the Zymogen form, aka inactive form of trypsin)
Within the intesntial cells there is a membrane bound enteropeptidase on the walls of the intenstines, which will cleave the N-terminus of the trypsinogen which then creates the functional trypsin
The workign trypsin then actiavtes other preoteolytic enzymes
The final step of the digestion of proteins are Amino acids
within the small intestens single, di, and tri peptides are transported into the blood stream via transporters on the basal side of the mucosa, while some proteins are digested by proteases and peptiadeses which are relased by the stomach, pancreas, and intestinal absorbative cells
Carbohydrate digestion
Carbohydrates produce 4 kcal/g and the energy comes from 70% startch and 30% sucrose
Carbohydrates digestion first starts in the mouth with a a-amaylase present within the salivia
this enzyme cleaves the a 1-4 glycosidic bonds and starts to digest startch and other polysccharides turning them into smaller units like glucose, maltose, and others
Carbohydrates travels to the stomach and small intenses to get more digested where the a-amylase and other enzymes digest various gylcosdiic bonds
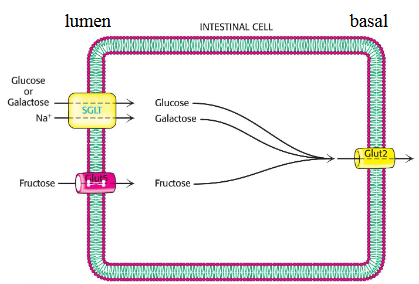
Sucrase lactase and maltase are present on the lumen side (the outside of the cell which faces the incoming food) of the cell membrane in the intestnial cells. These enzymes take their respective sugars and turn them into monosaccharides
glucose and galactose are then transported by the sodium glucose linked transporter whereas their counter part fructose is transported by Glut5
All the monoscharides are transported out of the cell by Glut2 which is on the basal membrane facing the blood vessels
Lipid Digestion
Triglycerides produce 9kcal/g which is roughly 30% of the daily caloric intake
Triglycerides are insobule in water and enzymes are solbule in water, so how do we digest these energy sources?
we emulsify the lipid making it able to be partially soluble in water which then lets the enzyme work in digesting it
to help emulsify these lipids, we use mechnaical motions like chewing, movments of the stomach and small intestines.
we have amphiathic emulsifers, such as bile salts, and phosppholipids
Lipases is a enzyme released by the pancreas which functions at the fat-water interface to hydrolyze the triglyceride in to free fatty acids and monoacylgrcerol
These FFA and MAG form what we call micelles, these lipid micelles diffuse into the intestinal cells while some are transported in by fatty acid binding proteins
FFA’s and MAg are then transported into the smooth endoplasmic reticulum of the interstinal cells which then they get resynthiszed into triclycerides
These newly resynthesized TG along with phospholipds, cholesterol, and apolipoproteins are packing into chylomicrons are then released in the lymph system
Chylomincrons are transported to many tissues where these TG are then trapsorted into cells, Chylomincrons also carry soluble fat vitmains like vitamin A and E
Food digestions have a role in nutrients and hormone
increase in glucouse within the blood stream triggers the response of the hormone inslunin, from pancreatic B-cells
insulin is a horomone which tells the liver, muscles, and other adipose tissues to uptake glucose, fatty acids, amino acids, which are molcules are used in anabolic pathaways.
After inuslin is released it tells the adipose tissues to rpocude leptin, which acts on its corrosponing receptors to represees orexingeric neurons and stimulate anoxrexigenic neurons, which in turns produces neurons to make the feeling of satiety
Low blood sugar glucose stiulates the glucagon production of pancreatic a-cells
glucaon is a horone in the liver, muscle, and adipose tissues to stimulate the catabolic pathway, this will breakdown fatty acids strored to help increase the glucouse within the blood.
In response ot a low glucose level in the blood some cells will release Ghrelin, which represses anorexigenric neurons and leads to hunger
Chapter 15 Metabolism
What is the purpose of metabolism within organisms and why is it needed to survive?
One of the reasons for metabolism is so that we can extract the energy from the food we eat and convert them into usuable forms of energy, this is called Bioenergestics
Another reason is to use the small molecules within the enviorment and within the food we eat to make our own macromolecules and their deritivatives to help our own body function
The pathway for metabolism is a forms of lines that connect and dissect at various points
which means you could start a one line for one product but then make a whole diffrent product
Metabolic pathways are connected steps that creat products that will be the reactants of the next reaction down the line
There are two diffrent catergories for metaboli pathways
Catabolic pathways
Anabolic pathways
Catabolic pathways are a series of reaction(s) that breakdwon a larger molecule into smaller parts to extract energy
EX: glycosis, lipolysis, Fatty acid oxidation, Amino Acid catabolism
Anabolic Pathways are series of reactions that use energy in order to create more complex biomolecules
Ex: Gluconegensis, Fatty acid syntehsis, lipogenesis
Oxidation of energy fules provide energy for Metabolism
Carbons that have lots of C-H bonds will provide lots of energy, however to extract that energy we oxidiate the molecule to release it attatching an Oxygen which is a stable molecule that will give us no energy
The act of converting biomolecules of C-H bonds into CO2 releases the energy we desire and thus drives metabolism
The oxidation fo energy fuel in catbolism happens in a stepwise manner
Catabolism generally happens the same way for fats, polysaccharides, and proteins
In the first step its the inital breaking down of the larger molecules into smaller molecules to be proccesed
this step uses energy rather than creating energy
The second step breaks down the smaller molecules into their simple units, usually generating Acetly CoA
This step generates a small ammount of ATP
In the third step we take the Acetyl CoA and oxidize it to generate a abundent ammount of ATP
Energy is stored in diffrent forms within metabolism
ATP is the main energy currency since its a high energy molecule
It has two phosphoanhydride bonds, and when undergoing a hydrolysis reaction it releases a large ammount of energy
ATP is als able to transfer its energy to other matabolites
This is called high phosphryl transfer potential
In the world of biomolecules the transfer of the phosphoryl group to a metabolite means the energy stored can have high affinty or low affinity, meaning if a molecule with high affinity gets tranfered a phosphoryl group from ATP, its able to produce more energy than norma ATP.
we call these a high phosphoryl transfer
an example of this is PEP, 1,3-BPG, P-Creatine
these molecules can generate ATP via Substrate level phosphorylation
Substrate level phosphorylation is a means of generating ATP by taking the energy from a high energy bond to make ADP turn into ATP
this happens from the generation of high eenergy phosphoanhydride bond between the beta and gamma phosphate groups
Metabolism turns the energy we can into five forms
Chemical energy
Chemical bonds
Kinetic Energy,
Movment
Potential energy
Proton gradient within the mithocndria
Electromagnetic energy
High energy elections in NADH and FADH2
Thermal Energy
Bondy temperature
Phosphorylation reacitons is when a phosphoryl transfer happens between ATP and a substrate
This is thermodynamically stable and Kinetically stable
meanign this has a high phosphoyl trasnfer potential and the transfer happens fast
This reaction is catalyzed by a enzyme called kinases
This type of reaction can happen on a protein, lipid, nucleotid, or carobhydrate
When this phosphrylation happens on a protein it occurs at the hydroxyl group in the amino acid residue side chain
Phosphorylation of a protein is very cruical as it plays a role in the regulation of metabolism and other cellular pathways
Amino acids residues in a protein that can be phosphorylated
Serine> Threonin> tyrosin and aspartate
When these residues are phsphorylated it changes the shape of the molecule and determines the enyzmes activaties and how it interacts with other molecules
ATP is the universal phsophryl donor and carrier for kinases
NAD+ and FAD are activated carrier of high energy electrons use in catabolism
NAD+ usually oxidizes alchols into aldehydes or ketones
FAD oxidizes lakanes into alkenes, saturated to unstaturated fatty acdis
WHen catabolism happens the breakage between the C-H produces high energy electrons that get trasnfered to NAD+ and FAD
In their reduced form of NADH and FADH2 at the end these electrosn get trasnfered to O2
Dehydrogenase is usualy hte enzyme that catalyzes the reactio ntrasnfer between NAD+ and FAD
In Anabolism the primary electro carrier is NADP+
When syntheiszing biomolecules electrons get trasnfered to NADPH to the smaller buiding blocks, this makes it so the reduced form of the smaller buidlign blocks is present and it can then store the energy via chemical bonds
Enzymes within Anabolism know to use NADP+ instead of NAD+, due to the extra phosphate group rpesnet in the ribose ring
COenzyme A (CoA) is a carrier for Acyly groups
Co-Enzyme A is a activated carrier of Acyl Groups in both fuyel oxidation and biosynthesis
this enzyme reactions with acyl grousps to form acyl-CoA that has a high energy thioester bond present
Energy is require to create the thioester bond
Breaking this thioester bond leads to the rlease of acyl groups and the energy stored in the bond 0
Chapter 16 Glycolysis
Glycolysis is a catabolic pathway which turns 6 carob hexoses monosaccharide to a 3- carbon pyruvate
This happens in the cytosol (cytoplasm)
Glycolysis works by taking oxidizing the monoscharides to make the energy stored into ATP and NADH
The oxidation of monosaccharides through glysosis dosnt require O2 isntead requiring to be oxidized by NAD+
There is two stages to glycolysis and ten steps split between the two stages
Stage 1 of glycolysis
for 1 molecule of glucose 2 mols of ATP are consumed in order to trap that 1 molecule of glucouse within the celll
the second part of stage 1 results in using two kinases to break a C-C bond to turn the gluouse into a 3C GA-3P and a 3C DHAP,
the kinases involved use a collective total of 2 ATP in this stage
from stage 1 we achieve 2x GA-3P
Stage 2 of glycolysis
this is the ATp harvest stage
with the new generate products of stage 1 we can generate 4 ATP molecules
When oxidizing the aldeyhyes (GA-3P) into pyruvate this step releases energy
Two kinase molecules will use the energy released to phosphorylate ADP to ATP
The energy trasnferd turns 4 NAD+ molecules into NADH
The two major goals of Glycolysis
Makes small building blocks and generate ATP
During both stages of glycolysis small molecule building blocks are made to support the lipids, nucleotides, proteins, and protein glycosylation
Stage 1: 1-5 Steps of Glyocolysis
1. Phosphoyltation (regulated step)
This step traps in a glucose molecule, by adding a phosphate to a glucose molecule
this means that the glucose molecule cant pass through GLUTs (transport molecule for glucouse)
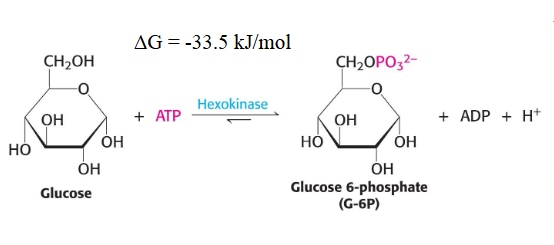
This is an irreversiable step since we used energy to create this which was catilizyed by hexokinase
HExokinase is regulated by the hypoxia inducible factor 1A and it is inhibited by the product its produces (glucose 6-P)
Kinases change their strucutre for their activity
Kinases need divalent cations, cations with a postive 2 charge, such as Mg and Ca
when the structure changes the glucouse is trapped within the enzyme and water is repelled away
this is becuase if water is present there is a chance the ATP hydrolysis without the phosphoryl trasnfer to glucouse, which means energy is wasted
2. Isomerization
This step turns the glucouse into fructose turning the 6 carbon ring into a 5 carbon ring, this will help the cleaving of a the 6 carbon sugar into a 5 carbon sugar
This step is cataluyzed by phosphoglucouse Isomerase (PGI) these catalyses the exchange of chemical groups on different carbons
3. Phoshprylation (regulated step)
The fructose molecule created in step 2 gets phosphrylated again and recieves a new phorphryl group at the 1-postion
This is a unstable molecule
this is catalyzed by a phosphofructokianse 1 (PFK1) and consumes ATP
This step is iireversible and is the commitment step, to which after this reaction occurs this molecule of fructose is destined to be cleaved and used for generating ATP
4. Adol cleaving
This step takes the unstable frutctose molecule created and cleaves it into two 3C triose phosphate glceraldehyde 3-phosphate(GA-3P) and a Dihydroxylacteon phsophate (DHAP)
when the two molecules are cleaved by the aldose enzyme the step is reversible
the bond broken is the 3-4 bond on the Fru 1,6-BP
5. Isomerization (a side step)
the DHAP molecule created in step 4 can be turned into a GA-3P molecule by the enzyme Triose Phosphate Isomerase
This makes it so the isomerization makes glycolysis as energy generation pathway
Stage 2 Steps 6-10
6. Phosphorylation Coupled to Oxidation
GAP is oxidized by removing the hydrogen off of the aldehyde end and replacing it with a phosphoryl group,
the hydrogen is attatched to a NAD+ group and is transfered by a GAP dehydrogenase enzyme
The reacttion first goes to a intermidate of a alchol group being attatched after being oxidized initally, this releases energy, however in order to add the phosphoryl group we have to kick off the alchol that was just added to replace it with a phosphoryl group
this means we must use energy in order to do it
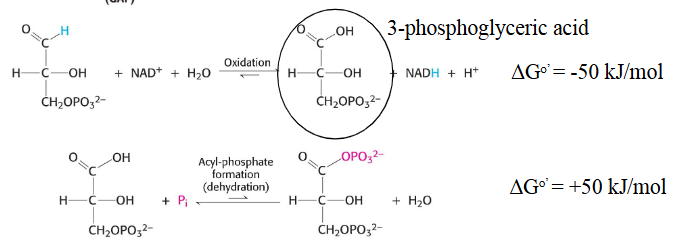
in order for this to not occur and we dont lose neergy in this process we have the enzyme and the intermdeiate formed create a thioester bond, which allows for a reaction that shouldnt work together happen
The take aways from this step is that GA 3-P is oxdized by the NAD+ we greated in an earlier step
The energy we release in this process goes into remaking NADH molecules and is used to increase phosphoryl transfer potential
1,3 BPG has a very high phosphoryl transfer potential that is sufficient to drive substrate level phosphorulation of ADP to ATP
7. Phosphoryl Transfer to ATP
Using the 1,3- biphosphoglycerate we are able to generate ATP via substrate level phosphorylatoin
This step is catalyzed by Phosphoglycerate Kinase, and is reversiable since it dosnt use much energy
This step generates one ATP
Once this step is completed the two ATP we have consumed in stage 1 is now returned back to us
8. Phosphoryl Shifting in position
This step takes the phosphoryl group that remains on the molecule and shifts it to the 2 position
This is done by a Phosphoglycerate Mutase (PGAM)
Mutase is usually an enzyme that takes chemical groups and shifts it intramoleculurly without distorting the shape of the molecule
9. Dehydration
This step takes water away from the molecule which in turn increases the phosphoryl trasnfer potential from 2-PG to PEP
This is very thermodynamically unfavorable, but since PEP is low in the cell the reaction is driven forward
10. Phosphoryl transfer to ATP (Regulated Step)
Using the PEP created in step 9, which has a High phosphoryl trasnfer we can generate ATP via substrate level phsophorylation
This is an irreversiable step as this requires energy
it is catalyzed by pyruvate kianase
many regualators in this step
Generates ATP
What is made through glycolysis
1 glucouse is turned into 2 pyruvate (3C) while 2 ATP and 2 NADH are generated
How does one maintain a fast glycolytic rate?
it requires a lot of NAD+,
this means we need a regeneration of NAD+ consistantly not to make new ones but to regnerate them once they become NADH+
Three ways to regenerate NAD+
1. Transfer the electrons gained from NADH and ship them off to the ETC within the mitochondria
this is used most of the time however its not that fast and has 3 preqreuistes
1. need abudnent ammount of O2
2. high output of the citric acid cycle
3. active transport of NAD+ and NADH in and out of the mitochondria
2. Take electrons from NADH and throw them into pyruvate to regenrate NAD+, it generates Lactate
This is a very fast regneration of NAD+ and is used in fast twitching muscle fibers during serious excerise
dosnt require oxygen!
3. Converting pyruvate into Acetaldehyde using a decarboxylatoin reaction
using the newly formed Acetaldehyde we take NADH and give it the electrons to form Ethanol
This is a process common in smaller organisms such as yeasts
Alchol Catabolism in humans
To detoxify alcohol we oxidize it to turn NADH into NAD+ and to turn the ethanol into Acetayl CoA
From the acetaldehyde, if we have too much in our blood to the point where our liver cant take care of it, it leaves to hang overs
Acetaldehyde is the toxic component in alchol metabolism
So it goes from Ethanol being converted into Acetaldehyde by Alcohol Dehydrogenase (ADH) then from Acetaldehyde it gets turned into Acetate by the enzyme Aldehyde dehydrogenase (ALDH). Then from Acetate it gets turned into Acetyl CoA, which guided by ACetyl-CoA synthetase and Acyyl-CoA Synthase 2
Regulation of Glycoylsis
There are three irreversiable steps within glycolysis
1. is when we turn glucose into glucose-6P
2. is when we take fructose 6P and turn it into fructose 1,6 BP which is catalyzed by phosphofructokinase 1 (PFK1)
3. is when we turn Phosphoenolpyruvate to Pyruvate which is catalyzed by pyruvate kinase
Regulation of Hexokinase
Hexokinase is inhibited by its own product due to allosteric inhibhibition.
This allows the cell to not use all the energy to turn Glucouse into glucouse 6-P all the time and just waste energy, since in times of rest we dont need to generate lots of ATP
Regulation of Phosphofructokinase 1 (PFK1)
PFK1 is repressed by Glucagon
This is because glucagon tells the cells to release glucose in order for it to be used in vital organs
however if we have PFK1 present we would be using this preciouse glucouse and it wouldnt make it to the cells that need it
PFK1 is allosterically activatyed by AMP and Fructose 2,6- biophosphate
this allows PFK1 to generate catalyze more reactions quicker to generate more ATP
Frucotse 2,6 Biophosphate also over turns the signal for ATP inhibition, meaning it tells the PFK1 to not stop and just to keep making ATP molecules, regardless of ATP present or not
PFK1 is allosterically inbihibited by
ATP inhibites PFK1 and is in competiation against AMP for the regulatory site
Phosphoenolpyruvate (PEP)
this is created in the later steps of glycolysis
Citrate
H+
Regulation of Pyruvate Kinase
To generate more Pyruvate Kinase PKL transcription is activated by carbohydrate responsie elemnt binding protein (ChREBP) and this protein is activated by G6P
this means when we eat and break down glucose we make more pyruvate kinase in order to get ready to store it
Pyruvate kinase is Allosterically actiavted by fructose 1,6-BP (PFK1 product)
Pyruvate kinase is allostercially inhibited buy ATP, Alanine (PKL but not PKM)
its also regulated by reversible covalent modifcations (Phosphorylation)
Fructose in Glycolysis
in Adipose/fat tissues the fructose gets turned into a fructose 6-P by using 1 ATP by hexokinase, it then just follows the regular steps that glucose would take in glycolysis
Fructose generates and uses the same ammount of energy as glucose
Within the liver cells resides a enzyme called Fructokinase which turns fructose into fructose 1-P by using 1 ATP molecule
Frucotse 1-P is Not a glycolysis intermediate
in the liver Fructose 1-P is cleaved into Glyceraldehyde and dihydroxyacetone phosphate by Fructose 1-P Adolase
Glyceraldehyde is then phophorylated by Triose kinase into glyceradleyde 3-Phosphate which also consumes 1 ATP
In the liver cells fructose by passes PFK1 and with this there is nothing to stop Fructose 1-P from being cleaved, which in turns makes Glyceraldehyde which is a toxic molecule and this will continously build up
Galactose in glycolysis
Making Galactose into glucose 1-P takes 4 reactions and 2 more ATP molecules used. Then it will take one more reaction to get it from Glucose 1-p to Glucose 6-P
This means when oxidizing Galactose it dosnt directly generate ATP, meaning that this is not a realible energy source for some cells
Cancers related to Glycolysis
Cancer cells will uptake Glucose faster than anyother cell
A F-FDG PET scan is used to find these cells, since F-FDG is a radioactive glucose, we can find the cancer cells which uptake it very fast
Tumor cells usually have a high activity of biosyntheis as well
In tumor tissues pyurvate will be converted to lactate under O2 rich conditions which is bad becuase its not suppose to happen, This effect is named Aerobic Glycoylsis or Warburg Effect
Chapter 17 Gluconeogensis
Energy Desnsities of Major energy fuels
Fatty Acids-9 Kcal Per gram
Glucsoe- 4Kcal per gram
Amino Acids- 4 Kcal Per gram
Ketone Bodies-
B-Hydroxybutyrate 5-Kcal pergram
Acetoacetate 4.15 Kcal Per gram
Acetone 7.4 Kcal per gram
During fasting the human body produces lots of Ketone bodies within the plasma of the blood on day 2 and it sky rockets per da. Glucose in the body goes down however it will level out as the days go on, since the body is taking stored energy and using it. Fatty acids are also insterted into the plasma at a higher rate but at a slightly higher level than normal.
Fatty acids are able to be used by a lot of tissues for an energy storach however when it comes to certain cells its not able to be used. For example the brain needs glucose, and since fatty acids cant be converted into glucose, we must have a constant balance of glucose within the body, in order to resume normal function
Why is Glucose still needed even in the fasting stages?
Glucose is needed in the red blood cells since it is their only source of energy
Since Red blood cells do not have a nucleus or mitochondria the metabolsim witihin these cells is simple
Within the brain is somthing called the Blood Brain Barrier, this is where red blood cells come and drop off nutrients for the brain
this is a highly selective barrier and only allows small molecules that are not polar to cross
Molecules that are able to pass the BBB and give the brain fuel is Glucouse, Amino Acids, and Ketone bodies, And creatine. All these can be carried by Blood
How is Glucose Produced During Fasting?
Glucogensis (GNG) is the process of of making glucose from Non Carbohydrate Precursos wtih at least three carbons
GLucose is not an essential nutrient for humans since it can be produced by our bodys
ONly two Tissues can prefrom this mkaking which are manly in the liver and kidneys
Glucogensis happens in the cytosol, mitcohondriam and the lumen of the smooth endoplasmic reticulum of cells
Glucogensis is activated after 2 days of fasting when, the last meals glucose supply is all used up
WHat is needed for Gluconeogensis
For anabolic reactions such as GNG we need precuros molecules to be joined together by new bonds, which involves adding electrons and energy into the reactoin
Substrates used in GNG
A lot of molecuels that contain at least 3 carbons can be used for the skeleton of Glucose
ATP generated from catbolic pathways are used
REductant: NADH is used and not NADPH
Acetyl CoA is not a precusor for GNG since it does not have enough carbons
For the energy required to produce GNG Fatty oxidation and Amino Acids Oxidation are required
THe precurosrs for GNG are
Pyruvate covnerted from Glucogenic amino acids
Pyruvate converted from Lactate
Oxalocaetate and other citric acid cycle intermediates
Glycerol
Cori Cycle
It is the process of the liver nad muscle tissues making and using glucose to support each others functions during a fast
The high energy electrons needed to process this is obtianed from glyocolysis within the muscles, then the electron is given to pyruvate which is later then turned into lactate
In Adipose tissues furing fasting Lipolysis is actiaved which realeases glycerol and Fatty acids from triglycerides
Both Glycerol and fatty acids are used in GNG
Fatty acids as a source of energy to power GNG
Glycerol as a precurosr for GNG
Glycerol entering GNG is diffrent from any other precursor
In the first step of it it reuqires ATP to be phosphorylated
the second step takes the newly formed Glceryol 3-phosphate and oxidizes 2 high energy electrons to form NADH and H+
The steps of Glucogensis
Glucogenesis cannot happen in the reverse steps of glycolysis since the reaction would be thermodynamically unfavorable
However Glucogensis does share most of the same steps glycolysis does except for a few exceptions
To make the three irreversible steps from glycolysis, reversible in glucogensis we use diffrent enzymes to make it thermodynamically favorable
For example:
Reaction converting PEP and pyruvate
glycolysis: it uses pyruvate kinase
Glucogensis: uses Pyruvate Carboxylase and PEP carboxykinase
Reaction Converting F1-6-BP and F-6P
Glucogensis: uses Fructose 1,6-Biophosphatase
Glycolysis: uses Phosphofructokinase 1
Reaction converting G-6P and Glucose
Glycolysis: Hexokinase/Glucokinase
Glucogensis: Glucose 6-Phosphatase
Step 1- Pyruvate to OAA (regulated)
Using Pyruvate Carboxylase i the mitochondral matrix of a cell Pyruvate is converted to Oxaloacetate
This is a regulated step by Acetyl-CoA, which activates Pyruvate Carboxylase
This step adds CO2 to the pyruvate which is adding a carbon and is the first step in commiting to making glucosee
The mechanism happens by Biotin Carboxylase transfering a phosphoryl group from ATP to bicarbonate to form a Caboxyphosphate
The biotin Prostehtic group in biotin Carboxyl carrier protein domain then catpures the activated Carboxyl group from phosphate as well as between bitonin and the BCCP
Lastly the BCCP then carries carboxyl bitoin enzyme strucutre and swing to the pyruvate carboxylase domain where the third step occurs where pyruvate is given another carbon
Step 1 occurs in the mitochondrial Matrix and is allostiercially activated by Acetyl CoA and allosterically inhibited by ADP
The ammount of pyruvate Carboxylase enzyme is induced by Glucagon and repressed by insulin
All Carboxylation reactions require a biotin prosthetic group as a carrier for actiavted CO2
Step 1.5- Transporting OAA out of the mitochondria
Within the Mtiochondra Oxaloacetate rises in concentration and with that the conversion of OAA and NADH to MAlate and NAD+ increases
Malate and high energy electrons are then transprotated out of the MItochondra
Step 2 OAA—→ PEP (regulated)
This reaction involves a decarboxylation and phosphorylation reaction at the same time
The energy that is utalized is the breaking of a C-C bond and GTP turning into GDP
This generates Phosphoenolpyruvate which is very unstable and high energy
This is a rate limiting step within Gluconeogensis
it is catalyzed by Phosphoenolypyuvate Carboxynkinase (PEPCK)
The ammount of PEPCK is induced by Glucagon and repressed by insulin
This is becuase we need PEPCK in fasted state to create more glucose however in a fed state we dont need to create glucose
ADP allosterically inhibits PEPCK
STEP 3-8 Common steps Between Glycolysis and GNG
Step 3: PEP—→ 2-Phosphoglycerate
Step 4: 2-Phosphoglycerate—> 3-Phosphogylcerate
Step 5: 3-Phosphoglycerate —→ 1,3- Phosphoglycerate
This step consumes 1 ATp to increase the phosphoryl transfer potential of 1,3 Biphosphaoglycerate
Step 6: 1,3 Biphosphoglycerate —> Glceraldehyde 3 phosphate
This is a redox reactio requiring NADH to reduce 1,3 BPG to GA3P
Step 7: glyceraldehyde 3-Phosphate—> Dihydroxyacetone Phosphate
Some Dihydroxyacetone Phosphates come from Glycerol
Step 8: GA3P + DHAP———> Fructose 1,6 Biphosphate
Step 9 (regulated): Fructose 1,6-BP——> Fructose 6-P
this step dosnt Generate ATP but instead takes the ester bond present and breaks it to relase energy, which makes this step exergonic and spontanous
This is catalyzed by Furctose 1,6-Biphosphatase, instead of PFK1 like in Glycolysis
Fructose 1,6 Biphosphatase is allosterically activated by Citrate
the opposite is true for PFK1, in glycolysis
Fructose 1,6 Biophostase is allostiercially inhibitied by AMP and Fructose 2,6biphosphate
The ammount of Fructose 1,6 Biphosphatase is also regulated by hormones glucagon and insulin
with lalot fo the PFK2 the enzyme turns the product it just made back into its starting material which is an inhibitor for Glycolysis which means glycolysis slows down and signals GNG to start
Step 11 (only applys to liver and kidney)
This is an irreversiabel step
Gucose 6-P is present on the SER of the liver cells and kidney cortex and the reaction hapenns inside the Lumen of the SER
Only liver cells and cells within the kidney cortex have G6P in their SER and so the glucose can be exported into the vesicles of the blood streams
Guconeogensis is very energy consuming
if the precursor is Lactate or Alanine
Costs 4 ATP, 2 GTP, and 2 NADH to make 1 glucose molecule
If the precurosors are oxaloacetate or aspartate,
Costs 2 ATP 2 GTP and 2 NADH to make 1 glucose molecuel
If precurosor is Glycerol
2 ATP and 2 NADH are required
GNG is very energy consuming and is only activated when the cellular energy charge is high in liver cells
Chapter 18- Pyruvate Dehydrogenase Complex
Glycolysis does not convert GLucose into Acetyl CoA
While Glycolysis happens in the cytosol and produces pyruvate, the pyruvate gets transferd to the mitochondral matrix where the enzyme Pyruvate dehydrogenase is located
Mitochondria Strucutre
The organelle is encolsed by two strucutres the, MOM and the MIM
Inner and outter membrane
Two comparments are found residing in the mithocondria
the Mitochondria matrix and the Intermembrane space
The MOM and MIM have diffrent permeabilites
The mitochondral matrix and the intermembrane have their own distinct pH’s and proteins/enzymes found present
The outer membrane has a higher permemability than the inner membrane
The outer membrane of the Mitochondria has Porin Complex on the membrane, which allows for free diffusion of most of the small biomolecules
Within the MIM is the Electron transport chain, ATP synthase, and some transporters for specific molecuels
Enzymes of oxidative metabolism are within the matrix
Pyruvate Dehydrogenase Complex (PDHC)
The complex has three subunits which makes this a very large protein strucutre
PDHC catalzyes catoblic reactions where the pyruvate loses a carbon (oxidation) and turns into an acyl group
PDHC extracts energy from the breaking of the C-C bond and uses that to generate high energy molecules such as
Acetyl CoA (Which has a high energy thioester bond)
NADH (High energy electrons)
PDHC does NOT generate ATP directly, but it helps in the process
PDHC catalyzes a series of biochemical reactions that turn pyruvate into Acetyl CoA
the order is Decarboxylation—> Oxidation——> Transfer to CoA
This uses three enzymes which are, pyruvate Dehydrogenase (E1), Dihydrolipoyl Transacetylase (E2), and Dihydrolipoul dehydrogenase (E3)
Pyruvate Dehydrogenase (E1) catalyzes the first step Decarboxylation and the oxidation of pyruvate
Thiamine Pyrophosphate (TPP) is a catalytic Cofactor in PDHc E1 Subunit
TPP has a carbanion which carries the acetyl group temporarly the acetyl group of the pyruvate
when TPP is carrying the pyruvate it becomes reduced and the carboxyl group in pyruvate is released as CO2
The first subreaction in E1 decarboxylates pyruvates
At the end of E1 when we have this new product, Lipomade from the next subunit E2 will swing itself into E1 and grab our product and take the acetyl group, off the product
This regenerates the TPP and the Lipomadde takes the acetyl group into the second sub unit
Lipoic acid is a catalyic cofactor in PDHc E2 subunit
Lipoamide brings the acetly group to Coenzyme A (a stoichiometirci cofactor of PDHC)
This forms Acetyl CoA
When the Acetyl group is ripped off the Lipoamide that was carrying it, it gets turned into the reduced form (Dihydrolipoamide) and this reduced form needs to be turned back into the lipoamide to continue more reactions
E3 Subunit ustilizes FAD to oxidize Dihydrolipoamide back to lipoamide and utilizes NAD+ to regenerate FAD
Within the E3 subunit FAD is used to take the two hydrogens off the reduced Lipoamide to regenerate it back into lipoamide
After this Lipoamide is made and FADH2 is made which is then oxidized by NAD+ to regenerate back into FAD
The high energy electrons that were orginally on Pyruvate is now on NADH
It first went from pyruvate, to Lipoamide, to FAD, to NAD+
There are Five Cofactors that are utalized in PDHc
Three are Catalytic cofactors
Thiamine Pyrophosphate (TPP) in E1
Lipoamide in E2
FAD in E3
Two are Stoichiometric cofactors (altered by reactions)
CoA which changes into Acetyl CoA in E2
NAD+ in E3 which changes into NADH as the last electron carrier
PDHc is tightly regualted
PDHc is tightyl regulated since it catalyzes a reaction that cannot be reversed in mammalian cells
this is becuase ACetyl CoA is not a precursor to Glucogensis
Once pyruvate is turned into Acetyl CoA it can be used in the Citric acid cycle or stored as a fatty acid
Since PDHC is a catabolic pathway which generates energy, it follows the same pattern as its other catabolic counterparts
Meaning that when it is in a higher energy state it is de regulated by allosteric inhibitors of its product it helps make such as aTp and NADH and Acetyl CoA
However when it is in a lower energy state molecules such as pyruvate and ADP allostercally activate it making it work more to produce more ammounts of ADP to get turned into ATP
E1 Subunits of PDHc is also regulated by Reversible Phosphorylation (catalyzed by PDKs and PDP)
Pyruvate dehydrogenase subunit can be phosphorylated by Pyruvate Dehydrogenase Kinases (PDK), which leads to a complete shut down of E1 activitey, which in turns means a inactive PDH
However when E1 is phosphoryalted it can be reversed by Pyruvate Dehydrogenase Phosphatase (PDP) to reactiavte PDH
PDK and PDP are allosterically regualted
PDKs are allosterically activated by ATP, NADH, and Acetyl CoA and allosterically inhibited by ADP, Pyruvate, CoA, and NAD+
PDP is allosterically activated by Ca2+ and Mg2+
Cancer metabolism and PDKS
Cancer cells will take pyruvate and convert it into lactate even when in O2 rich enviorments
this is becuase PDHc is not activated in many types of cancer cells, which leads to lactate being produced as the only energy source instead of ATP
in Cancer cells as well the gene HIF-1a is promoted which exhibits more PDK within cancer cells, which is the enzyme responsible for turning off PDHc
Chpater 19 Citric Acid Cycle (Krebs Cycle)
Just like the PDHc the Citric Acid cycle occurs in the mitochondrial matrix
There are three types of metabolic pathways
A liner pathway is when you take product A and go down a set of intermediates to achieve the final result
A Circular pathway, is using reactant A and then turning it into product F, but B, C ,D, E are regenerated to help produce F
A spiral pathway is used when lots of rectants AAAA, get used one by one until the last one A is standing
Cirtric Acid cycle
is a Catabolic Pathway
produces ATP
Acetyl Coa is oxidized by NAD+ and FAD and high energy electrons are extracted
Parts of the released energy is used to power substrate level ATP prodcution
Intermediates of the CAC
the intermediates of the CAC help the ACetyl CoA get oxidized
the helpers get regenerated so whether or not you have more Acetyl CoA it wont get processed without more helpers
2 Stages of CAC
Stage 1
Has 4 reactions where 1 Acetyl CoA (2C) is connected with a OAA (4C) and oxidized into 2 CO2 molecules
Stage 2
The last 4 reactions of the CAC where the 4C intermediate is transformed back into OAA
Both stages of the CAC generate high energy molecules like NADH, ATP, FADH2, with other anabolic pathways
CAC is a catabolic pathway designed to extract energy from Acetyl CoA
The chemical bonds within carbohydrates are mostyl oxdized by the citric acid cycle
this is becuase the stages before, Glycolysis and PDHc, are used to make the larger sugars into ACetyl CoA
Each turn of the CAC oxidizes one Acetyl CoA and generates
3 NADH
1 FADH2
1 ATP
Acetyl CoA is oxidized by NAD+ and FAD and NOT by oxygen
Step 1 of CAC (Cominbing OAA + ACetyl CoA and making Citrate)
OAA and Acetyl CoA combine to make a 6C
energy is released becuase we break the thioester bond in Acetyl CoA
This is a irreversiable step catalyzed by Citrate Synthase
For citrate synthase to work properly it is important that the loading sequance and induced fit work properly
Induced fit, means that the substrate has to be alinged perfectly for it to work and have the correct oriantation and alingment
Induced fit has two stages
1. the substrate OAA binds to the active side of the enzyme ( Citrate synthase) which allows active site to Induce fit the Acetyl CoA to create the intermediate Citryl CoA
2. After the creation of Citryl CoA the presence of the molecule indues conformational changes in the enzyme which breaks the Thioester bond in Citryl CoA
Usually Citrate Synthase wil not bind Acetyl CoA in its active site in order to minimize the chance of spotanous breaking of the thioester bond
Step 2- Turning Citrate into Isocitrate
This step involes dehydrating the molecule then hydrating the molecule to switch the alchol from position 3c to 2c
This is catalyzed by AConitase
the forward reaction of this is endergonic, this is becuase when isocitrate is created it gets consumed, which leads to a lower ammount of it which drives the reaction forward
Step 3 Isocitrate turns into a-Ketoglutarate (regulated)
This step is a REDOX reactio nwhich involves NAD+ coming in and taking 2 H atoms one from the alchol and one from the same carbon atom of the alchol
By oxidizing the C-O bond some of the energy gets stored in NADH
the result of breaking the C=O bond creates a intermediate called Oxalosuccinate which is highly unstable
The other part of the energy is used to break the C-C bond which contains the COO^- on carbon 3 which will be released as CO2
this reaciton is catalyzed by Isocitrate Dehydrgoenase in the mitochondria
The CO2 produced resutls for C1 of OAA not from Acetyl CoA
Step 4- a-Ketoglutarate turns into Succinyl CoA (regulated)
This is Oxadative decarboxylation reaction
This step extracts energy by Breaking the C-C bond, and storing some of the energy as NADH and another part of the energy is used to form a Thioester bond in the new product Succinyl CoA
This is highly regulated and also the RATE LIMITING STEP of the CAC
This reaction is catalzyed by A-KEtolgutarate Dehydrogenase Complex
Step 5- Succyinyl CoA turns into Succinate
In this step we break the thioester bond that we formed in step 4, and use the energy that is broken and turn it into ATP using substrate level phosphorylation
This is a thil lysis reaciton
This is a reversible step which is catalyzed by Succinyl CoA synthetase
this is the only reaction in CAC which generates ATP directly
Step 6- Succinate to Fumarate
This is a oxidizing step which takes away 2 hydrogens using FAD
FAD usualy is an oxidant for primarly hydrogens
the Electrons gained by FAD turn it into FADH2 and then the electrons get dumped to the Electron transport Chain
Siccinate Dehydrogenase is the major enzyme in Complex 2 of ETC
This step is an irreversible reaction in animal cells
it can be reversed in bacterial cells by using the enzyme Fumarate Reducatase thats found in bacteria
Step 7- Fumarate turns into Malate
This is a hydration step (adding H2O)
the Alkene function group gets added a Alchol and a hydrogen on one of the two carbons
This is a reversible step catalyzed by Fumarase
This is the connecting point with the Urea cycle
Step 8- Malate back to Oxaloacetate (OAA)
This a oxidizing reaction where NAD+ is the oxidant
This step extracts energy from the oxidation of the C-O bond turning it into a C=O bond
This regenerates OAA
This is a reversible step catalyzed by Malate Dehydrogenase
Both the forward and reverse reactions of this step are in GNG
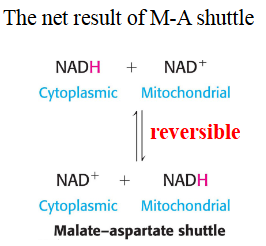
The forward and reverse steps are involved in Malate-ASpartate shuttle
used to shuttle electrons in NADH to go out or in the mitochondria
Within CAC Most of the energy released from glucose oxidation is stored in the form of NADH and FADH2, and not ATP
Regulation of the CAC- Steps 1,3,4,6
Cell Energy charge regulates the CAC flux
Higher energy molecules such as ATP, NADH, and Acetyl CoA allosterically inhibit CAC
In bacteria the higher energy molecuels ATP, NADH, ACetyl Coa allosterically inhibit citrate synthase (step 1 of CAC)
However all types of cells IDH3 (Isodehydrogenase 3) are allosterically activated by ADP and inhibited by ATP and NADH
In All cells a-KGDHc is allosterically inhibited by ATP and NADH and Succinyl CoA
When energy is high NADH and FADH2 is made alot which means that NAD+ and FAD supplies are low, which makes the CAC run slower
During starvation most of the OAA gets turned into Glucose via GNG and this process makes us feel like we dont have energy
CAC is connected with many other anabolic pathways to support biosynthesis
CAC intermediates are precurosrs for biosysntehsis
ACetly CoA is a precursor for fatty acids/sterols
to achieve this Acetyl CoA is brought out the matrix in the form of Citrate
a-Ketoglutrate and Oxaloacete are precurosors for amino acids and nucleotides
Succinyl CoA is a precursor for biosynthesis of Porphyrins,Heme, and Chloyprhyll
Oxaloacetate is a precursor for Glucose
Acetyl CoA cannot be converted into CAC intermediates
This is becuase Acetly CoA gets oxidized into two CO2 molecuels at the end of the COC
In order to get more CAC intermediates we can use anaplerotic reactions using nucleic acids, amino acids, and glucogenic amino acids to form new intermdeiates
Plants, Bacteria, and fungi can utilize Acetyl CoA to syntehsis glucose by short circuiting CAC- the Glyoxylate cycle
in lower organisms the presence of Isocitrate Lyase and Malate Synthase allows these organisms to go from Isocitrate to malate
this allows these organisms to use Acetyl CoA and condense it with Glyoxylate to form malate which can be turned into OAA
which supports the biosynthesis of glucose
Malate-Aspartate Shuttle and Glycerol 3-Phosphate shuttle
Malate Aspartate shuttle moves high energy electrons from Cytosolic NADH to mitochondrial NAD+
This is very active in the liver and heart and is used
This is a reversible shuttle
Glycerol 3-Phosphate Shuttle transports high energy electrons from Cytosolic NADH directly into ETC
High energy electrons are transfered from cytoslic NADH to FAD by two dehydrogenases
This dumps electrons at point Q directly, which means it is a very fast process and thus electrons cant be reveresed back on to DHCP
GLyceroyl shuttles are usually in the muscles since this is a fast acting shuttle it can be used to provide energy really fast
Defects in the CAC are fundamental causes of Cancers