Homoestasis and Cell Differentiation
Introduction to Physiology
Definition: Physiology is the study of the functions of organisms, encompassing various forms such as plant physiology, cell physiology, microbial physiology, and animal physiology. This text focuses on human physiology, examining how our bodies function. While emphasizing normal physiology, it occasionally describes pathophysiology to illustrate what occurs when normal body function is disrupted, using examples like diabetes to demonstrate the delicacy and interdependency of organ systems.
Systems Approach
Method: The systems approach studies one organ system at a time. An organ system is a collection of anatomical structures working together to perform a specific function. For example, the cardiovascular system delivers oxygen- and nutrient-rich blood to the body's organs. This approach highlights that no single system functions in isolation; systems interact, such as the urinary and cardiovascular systems.
Importance of Physiology
Curiosity: Understanding how the human body works is a common curiosity. Physiology is not just a collection of facts but a work in progress, with gaps in understanding and subject to change with new discoveries. Studying physiology broadens scientific outlooks, helping to see the "big picture" of body function as a connected whole. Many find beauty in physiology, making it their life's work.
Organization of the Body
Complexity: The human body is a complex, intricate structure with underlying simplicity in function. Despite its complexity, the body's operation can be explained by a small set of principles. For example, neurons, specialized to transmit electrical signals, illustrate this simplicity.
Interaction Among Body Parts
Cell Specialization: Cells are specialized to perform different functions essential for the body's operation. For instance, erythrocytes, produced by bone marrow, carry oxygen in the bloodstream. The synthesis of erythrocytes is regulated by erythropoietin, secreted by the kidneys. The heart's contractions, regulated by the nervous system, ensure adequate blood flow. The lungs, controlled by the nervous system, take in air to oxygenate the blood. The gastrointestinal system breaks down food into molecules absorbed into the bloodstream.
Cells, Tissues, Organs, and Organ Systems
Hierarchy: The human body is structured hierarchically, with cells forming tissues, tissues forming organs, and organs working together as organ systems.
Cells and Tissues
Cell Types: There are over 200 kinds of cells, grouped into four major classes: neurons, muscle cells, epithelial cells, and connective tissue cells. These classifications are based on functional differences.
Neurons: Specialized to transmit electrical signals, neurons have branches to receive or transmit signals. They enable perception, control of movement, hormone secretion, and cognitive functions.
Muscle Cells: Muscle fibers contract to generate force and movement. They are found in skeletal muscles (voluntary control), cardiac muscle (heart), and smooth muscle (blood vessels).
Epithelial Cells: Found in epithelia, these cells form barriers to separate body fluids from the external environment. They also transport materials and form glands (exocrine and endocrine).
Connective Tissue Cells: Diverse in structure and function, they provide support, anchor structures, and link parts of the body. Connective tissue includes blood, bone, and lymph.
Organs and Organ Systems
Organs: Structures composed of multiple tissues performing specific functions. For example, the heart pumps blood and contains muscle, nervous, epithelial, and connective tissues.
Organ Systems: Collections of organs working together to perform functions. Examples include the cardiovascular system (heart, blood vessels, blood) and the gastrointestinal system (mouth, stomach, intestines).
The Overall Body Plan: A Simplified View
Simplification: Physiologists simplify the body's complexity to highlight unifying themes. For example, the gastrointestinal system is depicted as a straight tube, and the heart as a box.
The Body's External and Internal Environments
External Environment: Epithelial tissue separates the external environment from the body's interior, including skin, lung linings, and gastrointestinal tract.
Internal Environment: Cells exchange materials with the bloodstream, not directly with the external environment. The internal environment includes the fluid surrounding cells and blood.
Exchange of Materials Between Environments
Lungs: Oxygen enters the bloodstream, and carbon dioxide exits during respiration.
Gastrointestinal Tract: Absorption of nutrients and secretion of digestive materials occur here.
Kidneys: Filtration, reabsorption, and secretion processes regulate fluid and waste.
Body Fluid Compartments
Water: The body contains water as a solvent for solutes. Compartments include intracellular fluid (ICF) and extracellular fluid (ECF), separated by cell membranes.
Homeostasis: A Central Organizing Principle of Physiology
Definition: Homeostasis maintains constant internal conditions despite external changes. It involves regulatory mechanisms across organ systems.
Regulation: Homeostasis regulates variables like body temperature and solute concentrations. Negative feedback mechanisms adjust variables to maintain set points.
Negative Feedback Control in Homeostasis
Mechanism: Negative feedback systems detect changes in regulated variables and initiate responses to counteract these changes, maintaining stability.
Example: Blood glucose regulation involves insulin secretion by the pancreas, reducing glucose levels through negative feedback.
Positive Feedback Systems
Mechanism: Positive feedback amplifies changes, useful in certain physiological processes like the LH surge triggering ovulation.
The Diabetes Epidemic
Definition: Diabetes mellitus affects blood glucose levels and impacts all body systems. It is characterized by excessive urine flow and thirst.
Prevalence and Impact
Statistics: Diabetes prevalence varies by age and ethnicity. It is increasing globally, with significant medical costs.
Obesity and Diabetes
Correlation: Obesity, defined by BMI, is a major factor in diabetes prevalence. Sedentary lifestyles and diets high in fats and carbohydrates contribute to obesity and diabetes.
Classification of Diabetes
Types: Diabetes includes type 1, type 2, gestational diabetes, and diabetes insipidus. Type 2 is most common, characterized by insulin resistance.
Diagnosing Diabetes Mellitus
Tests: Fasting plasma glucose and glucose tolerance tests diagnose diabetes. Symptoms include elevated blood glucose and glucose in urine.
Treatment of Diabetes Mellitus
Management: Treatment involves diet, monitoring blood glucose, and medications like insulin. Lifestyle changes and pharmaceutical interventions help manage the disease.
Cell Differentiation and Tissue
Within multicellular organisms, tissues are organized communities of cells that work together to carry out a specific function. The exact role of a tissue in an organism depends on what types of cells it contains. For example, the endothelial tissue that lines the human gastrointestinal tract consists of several cell types. Some of these cells absorb nutrients from the digestive contents, whereas others (called goblet cells) secrete a lubricating mucus that helps the contents travel smoothly.
However, the multiple cell types within a tissue don't just have different functions. They also have different transcriptional programs and may well divide at different rates. Proper regulation of these rates is essential to tissue maintenance and repair. The spatial organization of the cells that form a tissue is also central to the tissue's function and survival. This organization depends in part on polarity, or the orientation of particular cells in their place. Of course, external signals from neighboring cells or from the extracellular matrix are also important influences on the arrangement of cells in a tissue.
What Is the Source of New Cells for Tissues?
Without cell division, long-term tissue survival would be impossible. Inside every tissue, cells are constantly replenishing themselves through the process of division, although the rate of turnover may vary widely between different cell types in the same tissue. For example, in adult mammal brains, neurons rarely divide. However, glial cells in the brain continue to divide throughout a mammal's adult life. Mammalian epithelial cells also turn over regularly, typically every few days.
Neurons are not the only cells that lose their ability to divide as they mature. In fact, many differentiated cells lose this ability. To help counteract this loss, tissues maintain stem cells to serve as a reservoir of undifferentiated cells. Stem cells typically have the capacity to mature into many different cell types. Transcription factors — proteins that regulate which genes are transcribed in a cell — appear to be essential to determining the pathway particular stem cells take as they differentiate. For example, both intestinal absorptive cells and goblet cells arise from the same stem cell population, but divergent transcriptional programs cause them to mature into dramatically different cells (Figure 1).
Whenever stem cells are called upon to generate a particular type of cell, they undergo an asymmetric cell division. With asymmetric division, each of the two resulting daughter cells has its own unique life course. In this case, one of the daughter cells has a finite capacity for cell division and begins to differentiate, whereas the other daughter cell remains a stem cell with unlimited proliferative ability.
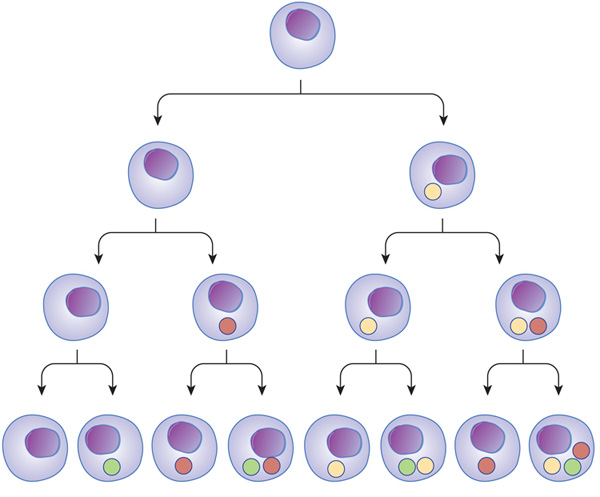
Figure 1: Transcriptional regulators can act at different stages, and in different combinations, through the path of cell development and differentiation.
Transcription factors can turn on at different times during cell differentiation. As cells mature and go through different stages (arrows), transcription factors (colored balls) can act on gene expression and change the cell in different ways. This change affects the next generation of cells derived from that cell. In subsequent generations, it is the combination of different transcription factors that can ultimately determine cell type.
How Do Non-Growing Tissues Maintain Themselves?
Although most of the tissues in adult organisms maintain a constant size, the cells that make up these tissues are constantly turning over. Therefore, in order for a particular tissue to stay the same size, its rates of cell death and cell division must remain in balance.
A variety of factors can trigger cell death in a tissue. For example, the process of apoptosis, or programmed cell death, selectively removes damaged cells — including those with DNA damage or defective mitochondria. During apoptosis, cellular proteases and nucleases are activated, and cells self-destruct. Cells also monitor the survival factors and negative signals they receive from other cells before initiating programmed cell death. Once apoptosis begins, it proceeds quickly, leaving behind small fragments with recognizable bits of the nuclear material. Specialized cells then rapidly ingest and degrade these fragments, making evidence of apoptosis difficult to detect.
What Cellular Components Support Tissue Structure?
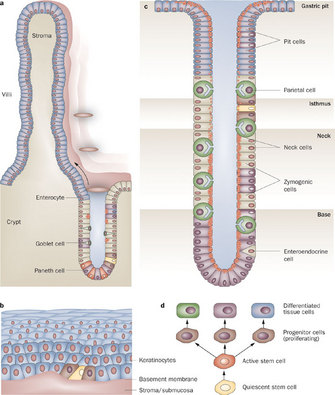
Figure 2 : Different cell types in the mammalian gut
The gut contains a mixture of differentiated cells and stem cells. The (a) intestine, (b) esophagus, and (c) stomach are shown. Through asymmetric division, quiescent stem cells (d) probably give rise to more rapidly dividing active stem cells, which then produce progenitor cells while losing their multipotency and ability to proliferate. All these progeny cells have defined positions in the different organs. To maintain its function and continue to produce new stem cells, a stem cell can also divide into and produce more stem cells at the same position (symmetric division).
Figure Detail
Tissue function depends on more than cell type and proper rates of death and division: It is also a function of cellular arrangement. Both cell junctions and cytoskeletal networks help stabilize tissue architecture. For instance, the cells that make up human epithelial tissue attach to one another through several types of adhesive junctions. Characteristic transmembrane proteins provide the basis for each of the different types of junctions. At these junctions, transmembrane proteins on one cell interact with similar transmembrane proteins on adjacent cells. Special adaptor proteins then connect the resulting assembly to the cytoskeleton of each cell. The many connections formed between junctions and cytoskeletal proteins effectively produces a network that extends over many cells, providing mechanical strength to the epithelium.
The gut endothelium — actually an epithelium that lines the inner surface of the digestive tract — is an excellent example of these structures at work. Here, tight junctions between cells form a seal that prevents even small molecules and ions from moving across the endothelium. As a result, the endothelial cells themselves are responsible for determining which molecules pass from the gut lumen into the surrounding tissues. Meanwhile, adherens junctions based on transmembrane cadherin proteins provide mechanical support to the endothelium. These junctions are reinforced by attachment to an extensive array of actin filaments that underlie the apical — or lumen-facing — membrane. These organized collections of actin filaments also extend into the microvilli, which are the tiny fingerlike projections that protrude from the apical membrane into the gut lumen and increase the surface area available for nutrient absorption. Additional mechanical support comes from desmosomes, which appear as plaque-like structures under the cell membrane, attached to intermediate filaments. In fact, desmosome-intermediate filament networks extend across multiple cells, giving the endothelium sheetlike properties. In addition, within the gut there are stem cells that guarantee a steady supply of new cells that contribute to the multiple cell types necessary for this complex structure to function properly (Figure 2).
How Does the Extracellular Matrix Support Tissue Structure?
The extracellular matrix (ECM) is also critical to tissue structure, because it provides attachment sites for cells and relays information about the spatial position of a cell. The ECM consists of a mixture of proteins and polysaccharides produced by the endoplasmic reticula and Golgi apparatuses of nearby cells. Once synthesized, these molecules move to the appropriate side of the cell — such as the basal or apical face — where they are secreted. Final organization of the ECM then takes place outside the cell.
To understand how the ECM works, consider the two very different sides of the gut endothelium. One side of this tissue faces the lumen, where it comes in contact with digested food. The other side attaches to a specialized ECM support structure called the basal lamina. The basal lamina is composed of collagen and laminin proteins, as well as various other macromolecules. On this side of the endothelium, adhesive junctions attach cells to the ECM. Transmembrane integrin proteins in the junctions bind components of the ECM and recruit signaling proteins to their cytoplasmic sides. From there, the signals travel to the nucleus of each cell.
Conclusion
Tissues are communities of cells that have functions beyond what any single cell type could accomplish. Healthy tissues require the proper mix of cells, and the cells within them must be oriented correctly and dividing at an appropriate rate. In order to coordinate their function, organization, and rates of death and division, the cells in a tissue are constantly processing and responding to signals from one another and from the ECM around them.
All of the cells within a complex multicellular organism such as a human being contain the same DNA; however, the body of such an organism is clearly composed of many different types of cells. What, then, makes a liver cell different from a skin or muscle cell? The answer lies in the way each cell deploys its genome. In other words, the particular combination of genes that are turned on (expressed) or turned off (repressed) dictates cellular morphology (shape) and function. This process of gene expression is regulated by cues from both within and outside cells, and the interplay between these cues and the genome affects essentially all processes that occur during embryonic development and adult life.
Do All Cells Really Contain the Same DNA?
Several lines of evidence support the proposal that all of the cells within a multicellular organism contain the same genome. For instance, although you started as a single cell with a half-genome from each parent, that single cell quickly divided and new cells began to differentiate, or become different from each other. While this process of differentiation established a wide variety of cell types (e.g., skin, liver, muscle, etc.), it was not accompanied by any permanent loss of genetic material. This is demonstrated by the fact that fully differentiated cell types are still capable, within the right environment, of giving rise to an entire new animal. This capability was first shown by way of an experiment in which the nucleus of an adult frog skin cell was transplanted into an enucleated donor embryo, eventually leading to the development of a cloned adult frog (Gurdon et al., 1975). Later, the intact complete genome of a differentiated cell was used in the cloning of the famous sheep Dolly (Figure 1), showing that in mammals, genes are not lost during development, so they must therefore be regulated (Wilmut et al., 1997).
Today, researchers understand that the specialized, differentiated cell types of the adult body contain a genome as complete as any embryo's. This fascinating demonstration has led to the proposal that changes in gene expression, rather than losses of genetic material, play a key role in guiding and maintaining cell differentiation.
Cell-Extrinsic Regulation of Gene Expression
Gene expression is regulated by factors both extrinsic and intrinsic to the cell. Cell-extrinsic factors that regulate expression include environmental cues, such as small molecules, secreted proteins, temperature, and oxygen. These cues can originate from other cells within the organism, or they can come from the organism's environment. Within the organism, cells communicate with each other by sending and receiving secreted proteins, also known as growth factors, morphogens, cytokines, or signaling molecules. Receipt of these signaling molecules triggers intercellular signaling cascades that ultimately cause semipermanent changes in transcription or expression of genes. Such changes in gene expression can include turning genes completely on or off, or just slightly tweaking the level of transcript produced. This process is thought to regulate a vast number of cell behaviors, including cell fate decisions during embryogenesis, cell function, and chemotaxis.
In addition, gene expression changes can lead to changes in an entire organism, such as molting in insects. In Drosophila, for example, the molting process is regulated by levels of a hormone called ecdysone. This hormone acts as a signal, triggering a cascade of events and leading to changes in gene expression. Not surprisingly, the genes that are expressed in response to ecdysone are also the genes that are involved in the molting process (White et al., 1997). Thus, ecdysone acts on the organism level as a cell-extrinsic factor to bring about physiologically meaningful changes in gene expression.
What is also interesting is that scientists can learn more about a physiological process like metamorphosis by studying how gene expression patterns change over time. For example, although researchers were aware that ecdysone results in a decrease of transcription from some loci, such as those involved in the glycolytic pathway, microarray data suggest that ecdysone-induced metamorphosis also downregulates genes involved with fatty acid oxidation, amino acid metabolism, oxidative phosphorylation, and other pathways. This suggests that there is a more global repression of metabolic activity during molting (Figure 2). Specifically, during metamorphosis, the larval muscle cells are degraded, and muscle-specific genes are downregulated (Figure 2B). Simultaneously, the development of the nervous system begins, and the genes involved in neuronal differentiation are induced (Figure 2C).
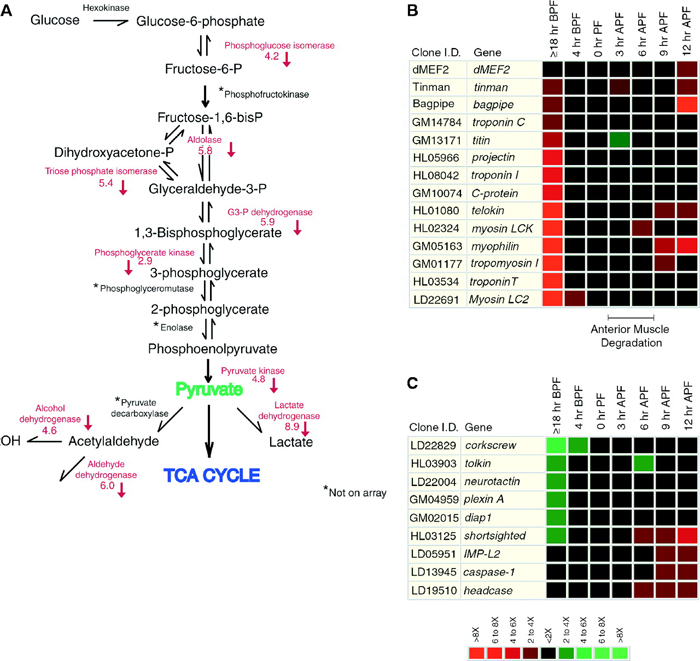
Figure 2: Microarray data collected at different times during metamorphosis reveals the effects of the ecdysone pulse on many downstream genetic pathways.
(A) Changes in ecdysone levels affect the glycolytic pathway. Levels of a number of enzymes involved in this pathway are decreased as a result of the ecdysone pulse; these enzymes are listed in red next to the reactions they catalyze. (B) This array shows expression changes in various structural and regulatory genes involved in muscle formation (myogenesis) in response to the ecdysone pulse. (C) This array shows how the ecdysone pulse alters expression of multiple genes involved in central nervous system restructuring, apoptosis, and cellular differentiation during metamorphosis. In both of the microarrays, red means that the gene was downregulated, while green means that the gene was upregulated. Expression levels were measured at various points before and after pupal formation (PF). (BPF = before pupal formation; APF = after pupal formation)
Cell-Intrinsic Regulation of Gene Expression
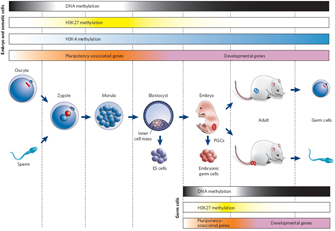
Figure 3: Epigenetic gene regulation during mammalian development.
© 2007 Nature Publishing Group Reik, W. Stability and flexibility of epigenetic gene regulation in mammalian development. Nature 447, 426 (2007). All rights reserved.
Figure Detail
Although differentiation is not thought to occur by permanent loss of genetic material, DNA can be modified in a way that affects gene expression. For instance, DNA and its associated histone proteins (together known as chromatin) can be chemically modified by a cell's own machinery. Chromatin modification can affect gene expression by changing the accessibility of genes to transcription factors, in either a positive or a negative manner. Two major classes of such chemical modifications include DNA methylation and histone modification (methylation and/or acetylation). These changes are often described as epigenetic because they do not act to alter the primary DNA sequence but instead act at a level just above the DNA sequence. Although DNA methylation and histone modification are not genetic, cells have mechanisms to copy this epigenetic information during their division so that their daughter cells contain the same regulatory data.
Changes in chromatin modification play an important role in regulating gene expression during developmental cell-type specification as well. For example, chromatin-modifying proteins play an essential role in muscle cell differentiation via interactions with key muscle-promoting transcription factors MyoD and MEF. That is, these factors are thought to help recruit chromatin modifying factors, such as histone acetyltransferases and deacetylases. In so doing, MyoD and MEF alter access to their target sites upstream of muscle differentiation genes. For instance, MyoD binds histone acetyltransferases p300 and PCAF, and this activity is essential for muscle cell differentiation (Puri et al., 1997). This example provides evidence for a link among chromatin modifications, transcription factors, and, ultimately, cell-fate-specific changes in gene expression.
Chromatin modification can be stable over the life of an organism, thereby effectively permanently influencing gene expression. However, that is not to say that chromatin modification is irreversible. For instance, chromatin can become mismodified in certain cancers (Vucic et al., 2008), suggesting that, although important, the change is not permanent. Moreover, chromatin modifications are usually erased and reset during the production of gametes, such that the adult program of intrinsic cues is replaced with a program more suited to embryonic development (Figure 3; Reik, 2007).
In fact, embryonic cell types are known to contain a unique set of chromatin modifications that are different from those found in adult cell types (Bernstein et al., 2006; Meshorer et al., 2006). This has led to the tantalizing proposal that chromatin modification helps lock in changes in gene expression that are required during development. The permanent silencing of the genes involved only in embryogenesis could then drive the development of cells toward more mature cell types. By blocking accessibility of transcription machinery, for example, chromatin modification could prevent the need for continued repression through active binding of a repressive transcription factor. Alternatively, the genes required for an adult cell type might contain chromatin modifications (especially histone acetylation) that cause the DNA to become open and, therefore, more accessible to the transcription machinery.
Interestingly, embryonic cell types have been found to contain a signature chromatin modification in the regions that regulate the expression of genes involved in early embryonic development (Bernstein et al., 2006). Such regions were found to contain chromatin modifications with both silencing and promoting characteristics. The finding of these bivalent (two-directional) markers in association with genes important for embryonic development has led to the belief that embryonic cells exist in a special epigenetic state, wherein they can choose to remain embryonic (as in an embryonic stem cell) or to differentiate (as in normal development), and bivalent domains provide a means by which to quickly choose between the two options.
Together, these lines of evidence have led to an emerging hypothesis that cell-cell signaling and epigenetic changes converge to guide cell differentiation decisions both during development and beyond.