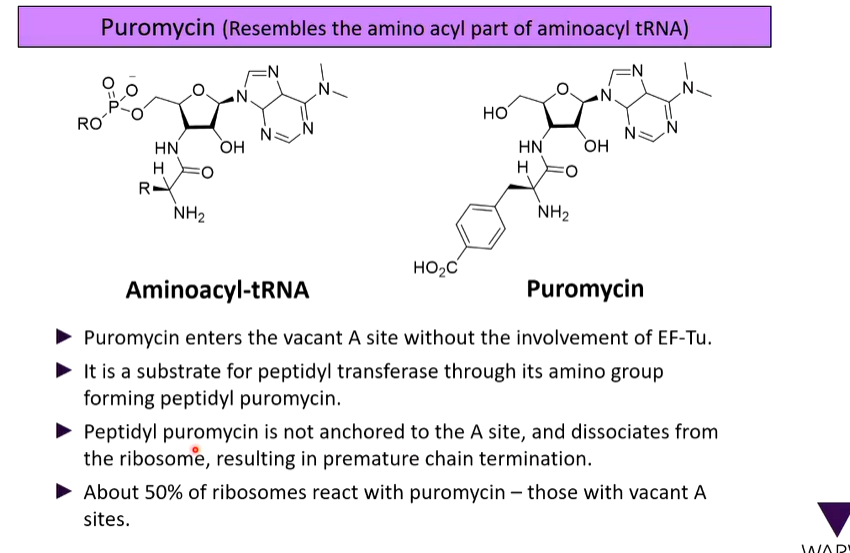
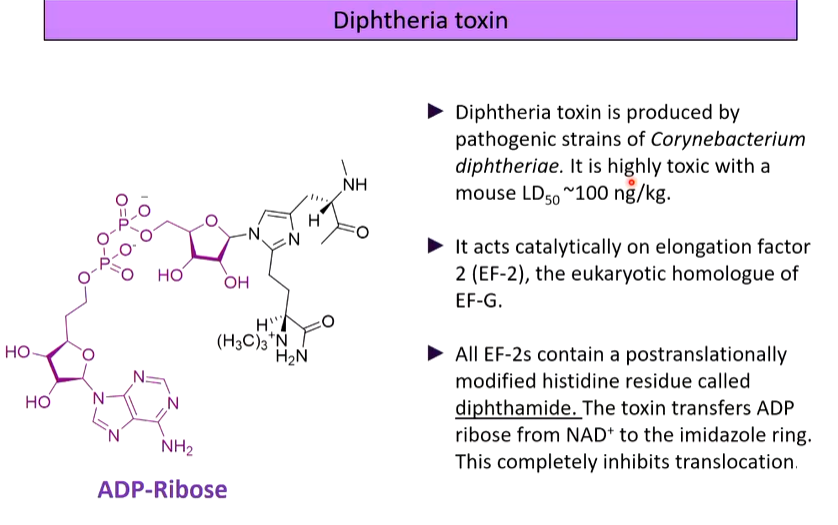
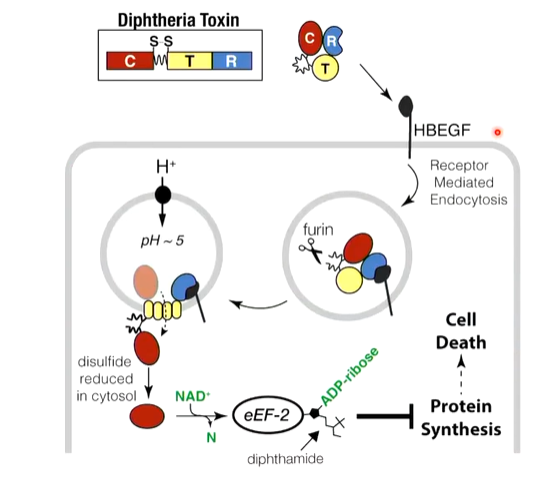
lecture 8: translation and translational control
Ribosome
structure
There are similarities between pro and eukaryotic ribosomes. In both cases, there is a larger subunit (60S for us and 50S for prokaryotes) and a smaller subunit (40s us and 30s them) to make the assembled ribosomes 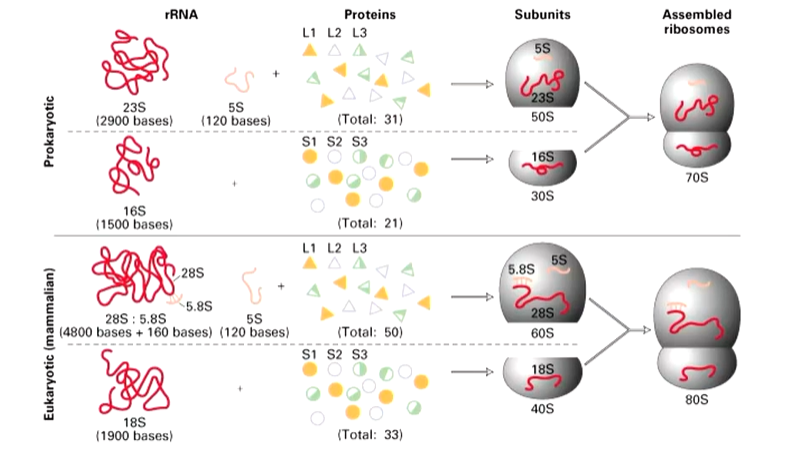
Each of the two parts is associated with a large number of proteins.
The large subunit is about 64000 atoms and the small unit is 29000 in prokaryotes.
rRNAs of e.coli were first sequenced in 1978 and secondary structure models were proposed based on base pairing. rRNAs from several hundred species have now been sequenced and all show the same defined structures - certain regions are always conserved. Mutations in these regions are lethal. There are variable regions as well which can tolerate mutations. There are compensating base changes between species and base paired stems (a forming helix) are common. 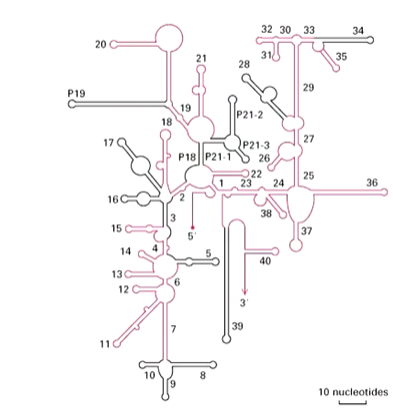
In the 3D structure, the superfolding is done by the loops done in the correct places. 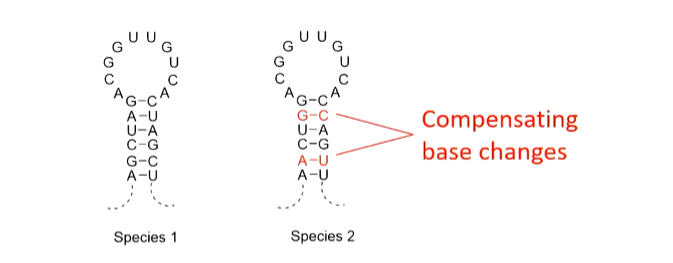
This shows the importance of the helical stem structure
chemistry
Ribosomes have three binding sites for tRNAs that span the 30S and 50S subunits. A is the acceptor site of codon directed binding of incoming aa tRNA. P is the peptidyl site that holds the codon directed peptidyl tRNA and E is the exit site that ks not associated with mRNA. These pockets are formed when the two subunits come together.
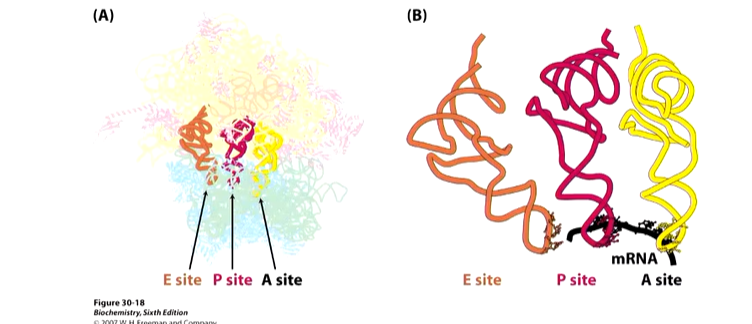
The reaction that happens in the ribosome is the formation of a new peptide bond. 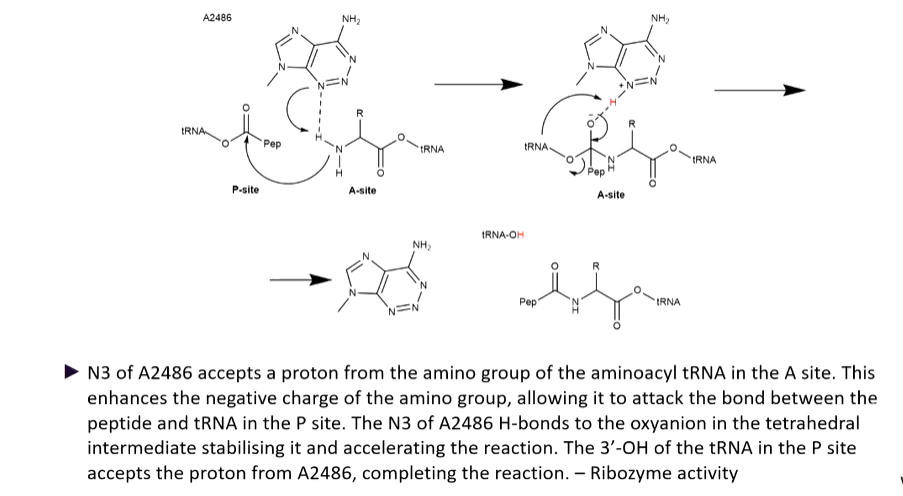
The attack of electrons come off the rRNA off the ribosome itself. this is ribozyme activity. The peptidyl transferase active site is associated with proteins where the closest one is 18 angstroms away - too far away to have any activity so is important for stabilization.
The protein goes through the exit tunnel as the protein is formed and this tunnel has a lot of hydrophobic residues which allows the formed protein to start folding with alpha helices fully formed when they come out. The protein itself is part of the mechanism for keeping the two parts of the ribosome together. there is a risk of premature stopping of the translation process which is prevented by the first amino acids that have been translated acting as a glue to keep the small and large subunits together. 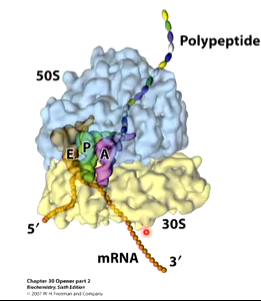
tRNAs
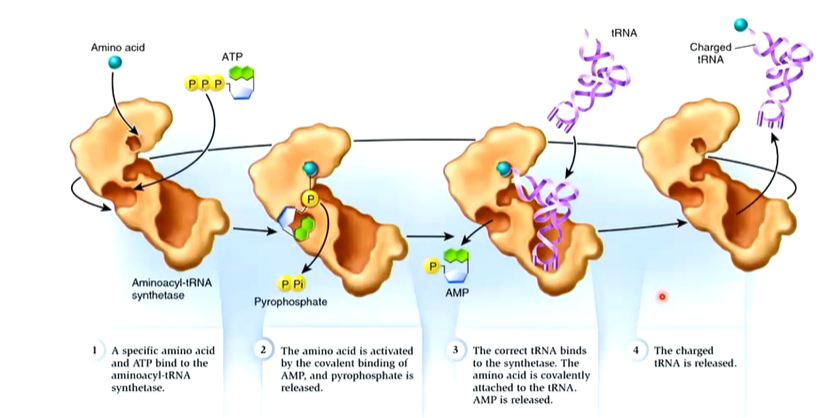
tRNAs are named after the amino acid they recognize and by to by an ester bond ege tRNAala. It is common for several tRNAs to become charged with the same amino acid and these are called isoaccepting tRNAs. The aminoacyl-tRNA synthetases (Enzyme which charge tRNAs - right amino acid on tright tRNA) show specificity for the tRNAs they charge and correct interaction is with cognate tRNAs. Rarely, non cognate (incorrect) tRNA is aminoacylated, tRNA contain a number of modified nucleosides which are unusual: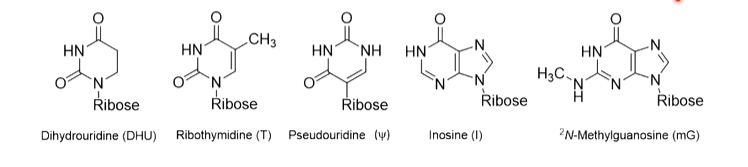
These are part of maintaining the 3D shape of the tRNAs as they have to be identical to each other to fit into ribosome pockets but also have enough differences to recognise the coding region and amino acid they are attached to
structure
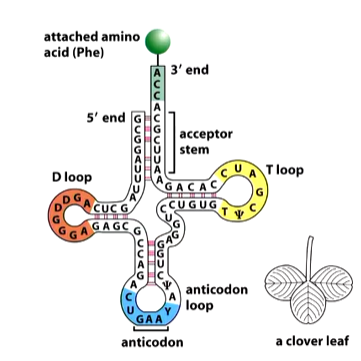
D loops contain 8-12 unpaired bases and contains 2-3 dihydrouricil residues
the anticodon loop is of 7 unpaired bases and contains the three anticodon bases. The codon is flanked on the 5’ side by U and on the 3’ site by an alkylated purine
the T loops is 7 unpaired bases and is involved in binding the tRNA to the a site of the ribosome.
The 3’ end always as CCAA amino acid attachment - the 4th base is variable and this is where the amino acid is attached.
The stems between loops are what give structure and are closely controlled sizes.
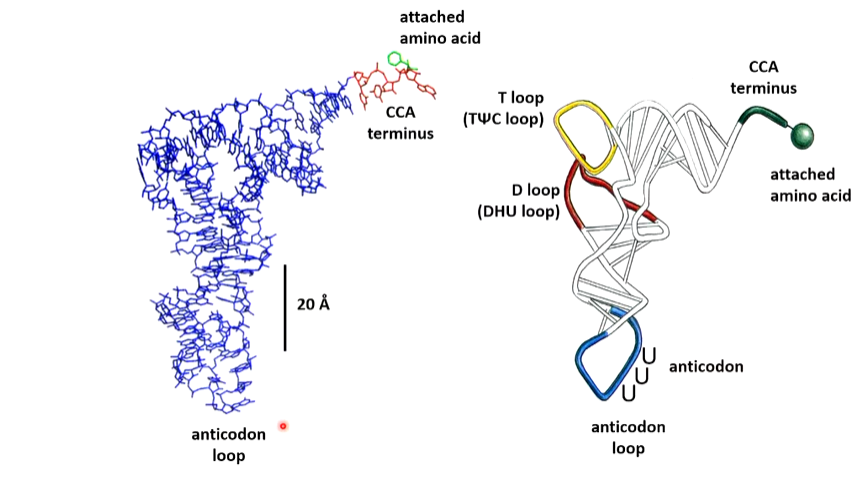
The CCA-3’ is located 70 A away form the anticodon the D and T loops form the L shape. Most of the bases are stacked which is a major factor in stabilization. The 3 anticodon bases and the CCA3’ bases ae unstacked which allows the interaction with the codon base or the aminoacyl tRNA synthetase
Many of the tertiary H bonding interaction involve bases that are invariant in all known tRNAs which strongly supports the belief that all tRNAs have the same structure. Many of these H bonds involve non conventional A-U and G-C pairs
shared and unique reactions of tRNAs

tRNA synthetases
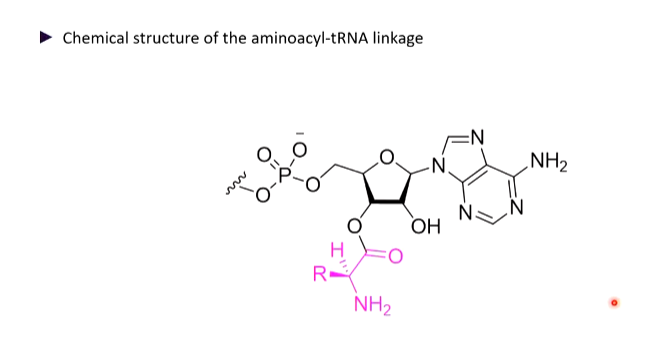
Ester link between aa and tRNA
Each synthetase recognises a single aa and all of the tRNAs that should be charged by it are called cognate tRNAs, Cognate tRNAs may differ in their anticodons and in other parts of the molecule. Aminoacyl tRNA synthetases are a diverse group of enzymes that have other functions
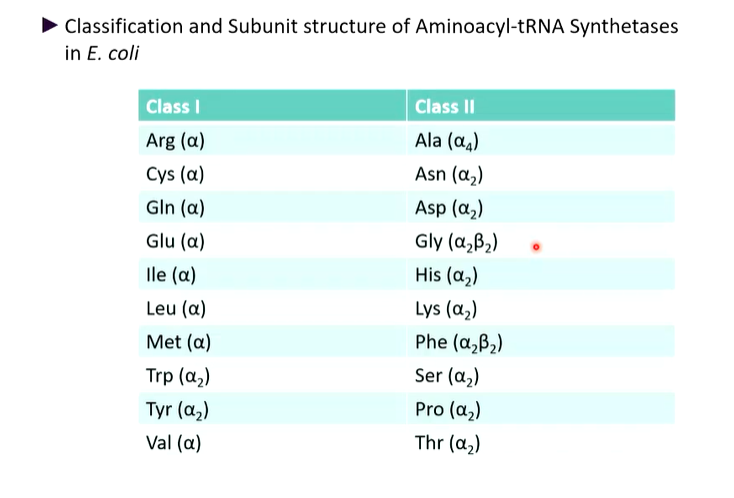
There are two general groups containing 10 enzymes:
Class I
contacts tRNA at a minor groove of the acceptor stem and anticodon
N terminal: a-b-a-b-
Class II
contacts tRNA at the major groove of the acceptor stem and anticodon.
beta surrounded by -a
Class I and II synthetases recognise different faces of the tRNA molecule. the CCA arm adopts different conformations 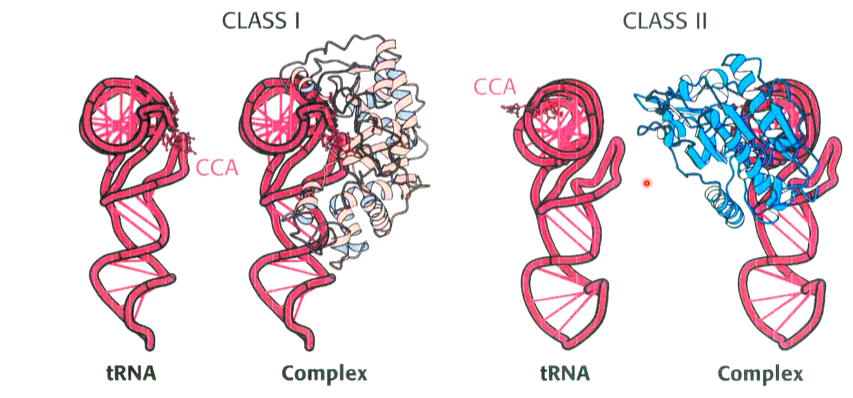
The process of translation is costly and it is important that the process is accurate. the features of individual tRNAs which re recognised by their cognate synthetase are called identity elements. They are on the anticodon bases of some tRNAs however for others, this is elsewhere. The significance of this is apparent from crystal structures of tRNA synthetase complexes
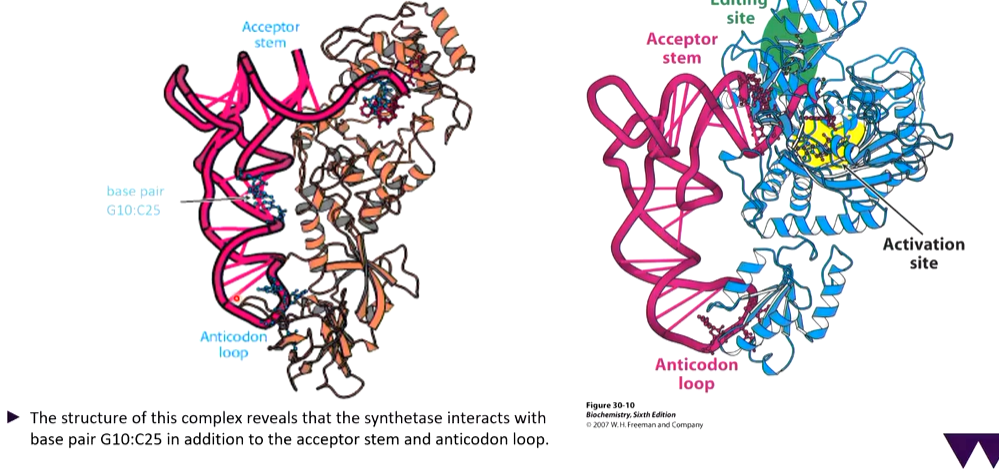
There are interactions in the bend of the L shape.
Threonyl-tRNA synthetase complex and its active site
The way the enzymes work is there is an amino acid that attaches to the enzyme and if this fits in the activation pockets, it sits there. If there is attachment of the aa on the CCA chain, this part moves between the activation and editing site. if it fits better in the editing site, the aa is cleaved off.
Proofreading can occur at two stages:
by hydrolysis of the ester bond of an incorrect aminoacyl-AMP intermediate triggered by the binding of the cognate tRNA
by hydrolysis of the ester bond of a miss matched aminoacyl tRNA
Most aminoacyl-tRNA synthetases process editing sites in addition to the acylation site and usually the acylation site rejects an amino acid larger than the cognate aa due to insufficient room. the editing site hydrolysis aminoacyl tRNAs which are smaller than the cognate aa.
How translation is used between species

Antibiotics target sites of actions in translation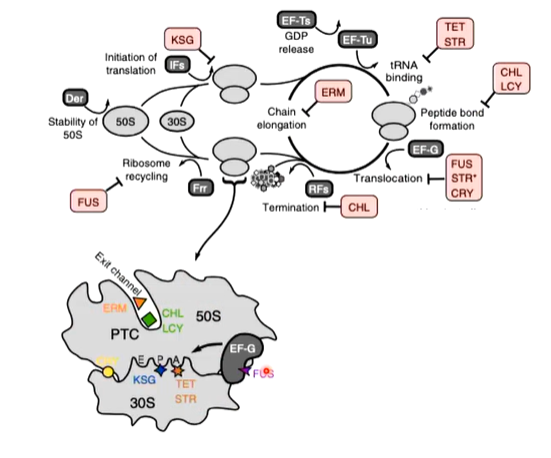

Streptomycin is a highly basic trisaccharide and binds to the 16s rRNA of the 30s subunit of the bacterial ribosome which interferes with the binding of formylmethionyl-tRNA to ribosomes which prevents the correct initiation of protein synthesis