lecture 5: transcriptional regulation
Chromatin structure
Transcriptional regulation is required as it:
allows development of different tissues
Transition from children to adult
deregulation can result in uncontrolled cell growth
allows reaction to environmental cues
chromatin structure, RNA polymerase binding specificity and additional binding and activation factors determines how/when and why genes are transcribed
Histone code revisited
The histone code is what determines whether the structure is open or closed. This shifts the balance between expression and silencing. Genes within highly packed heterochromatin are usually not expressed.
On histone H3, methylation on position 9 on lysine is usually associated with gene silencing, however the type of heterochromatin formed is dependent on if it is tri or uni methylated.
Inactivation of the HOX genes can occur by methylation of the lysine at position 27 which leads to x chromosome inactivation. Hox genes are developmental genes that are arrayed in the same order as the segments of the body. To ensure the right genes are expressed in the right segment, there is wholesale closing of whole chunks of HOX genes through this epigenetic mark. Two x chromosomes means that there is the danger of gene overdose.
On the H3 chromosome, acetylation of lysine at position K 14 + phosphorylation at S 10 and methylation at K 4 + K9 both lead to increased gene expression. The methylation at K4 is a single to make the DNA stay open.. This is important for the structure of the centromere which have to have an open conformation at all times
This methylation and acetylation bind proteins which are what regulate structure
structure of an interphase chromosome
Chromatin is mobile - the 30nm fibre is compacted into loops which is held together by scaffold proteins which has a lot of totoisomerases - implies a lot of winding and unwinding. It was though that loops can extend where they need to be expressed and there is some evidence to support this: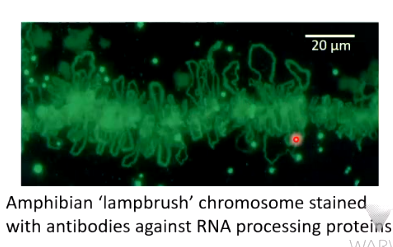
Chromosomes stained for transcriptional activity show there are actively transcribed loops
Chromatin is known to move within the nucleus: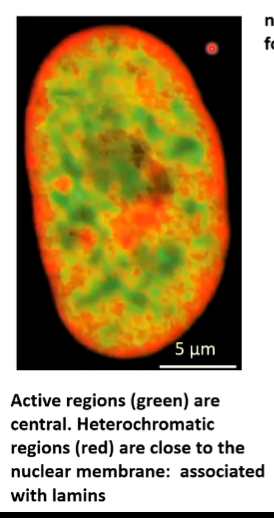
Red areas are not transcribed and are around the edges of the nucleus. This is where DNA repairs take place and heterochromatin formation. If there is a gene that needs to be turned on, the loop is moved into an area where the requirements for transcription are concentrated. If it needs to be shut down, it is moved to nucleus periphery where enzymes associated with the lamina shut it down.
There are two broad types of heterochromatin: 
facultative - gene that is previously on can be turned off and can be turned into euchromatin if it needs to be turned back on.
constitutive - consistently silenced. Areas can spread to make sure gene expression around them are also silenced
Human centromere organisation
A human centromere is made up of a centric heterochromatin which is long and highly repetitive chromatin structures. This is a set of nucleosomes with the normal H3 that is double methylated at K 4. This is on either side of centromere specific H3. Beside this, the centric is flanked by the pericentric heterochromatin.
The reason for this is that the centromeric histone forces an open structure and the key double methylation also does this. This means that centromeres remain open which allows access to all the kinetochore proteins that associate with centromeres which is the focus point where two chromatids are taken apart
Telomeres
Telomeres vary in length and occur at the end of chromosomes with the structure identifies in tetrahymena. It is structurally a set of thousands of repeat units (GGGTTG) but in ours the 3’ G is replaced with an A (GGGTTA). In yeast, this is flexible but function the same way.
RECAP END REPLICATION PROBLEM 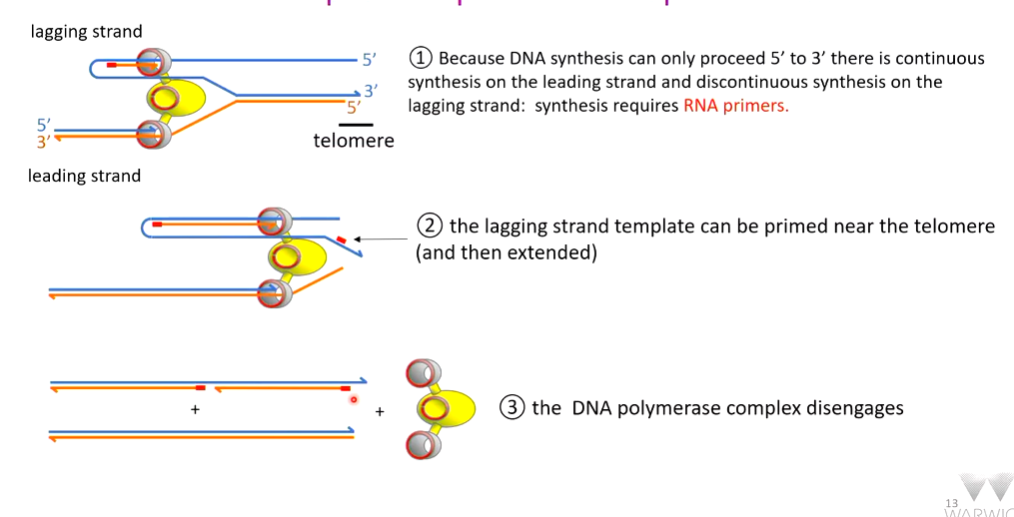

There is a compensatory mechanism for telomere shortening. This is done by an enzyme called TERT (telomerase reverse transcriptase). Telomerase is made up of 4 major protein romains with a long RNA running through it which holds together and stabilizes the 4 domains. Part of this RNA sequence has an AC which can hybridise with the TG terminal bases on the parental strand. The (AC)CCCAAC sequence acts as a template to make new DNA that is extended from the 3’ end. The telomerase repeats this - RNA directed DNA synthesis. This partial lengthening of the telomeres provides more room for DNA primase to lay down an RNA primer which has a 3’ hydroxyl that can be extended by DNA polymerase to repair part of the frayed telomere.
There is still a free 3’ unpaired end and this normally triggers DNA repair mechanisms which if triggered during mitosis telomeres will fuse. This doesn’t normally happen so the 3’ end shelters from repair mechanisms:
sheltering of the 3’ end
The 3’ end is sheltered in a complex called shelterin which is a complex of:
TRF1 - telomeric repeat binding factor 1
TRF 2 - telomeric repeat binding factor 2
RAP 1 - repressor/activator protein
There is also a 3’ overhang on the parental strand. The DNA strand is associated with the shelterin complex. These proteins protect the telomeric end with the reactive 3’ end coiling back to invade the ds DNA that has the same repetitive structure and displaces a D loop. The 3’ end is now base paired and no longer a danger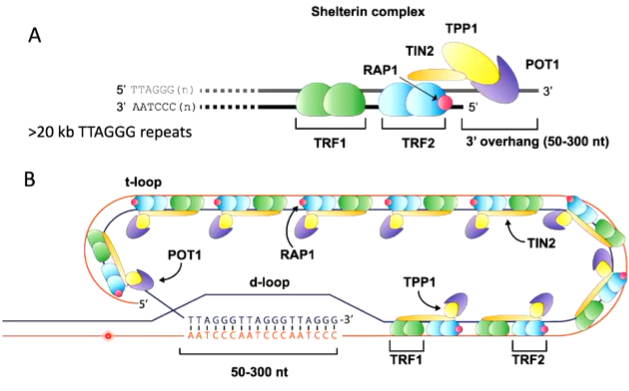
When this goes wrong, enhanced aging such as Werner’s syndrome occurs. The WRN helicase protein is important for DNA repair and telomeric DNA replication. Telomeres that are normally replicated by lagging strand synthesis are not replicated efficiently in werner cells. overexpression of telomerase in vitro counteracts with WRN mutation
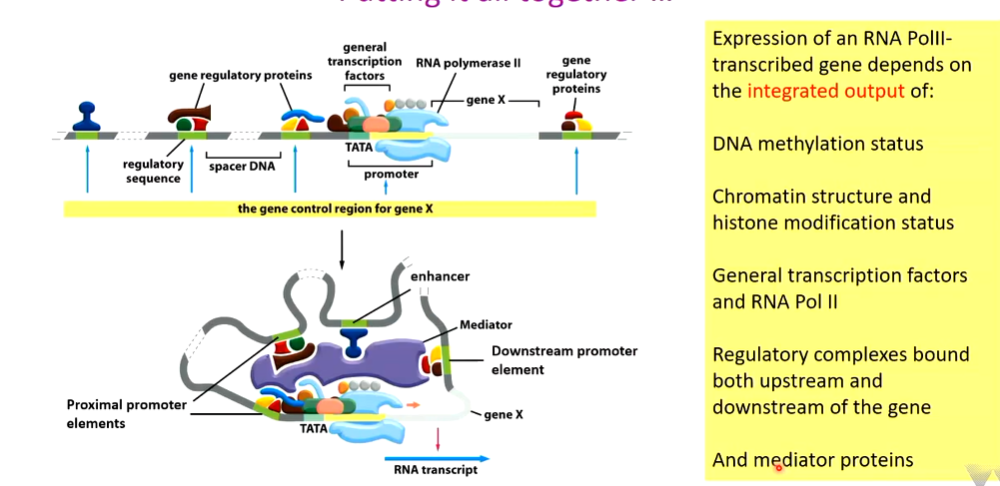
RNA polymerase and general transcription factor binding
transcription can also be controlled by RNA polymerase and general transcription factor binding specificity.
There are three major RNA polymerases. They only make up 0.0001% of total cell protein.
RNA pol II is focused here.
Human Pol II promoter looks like this: 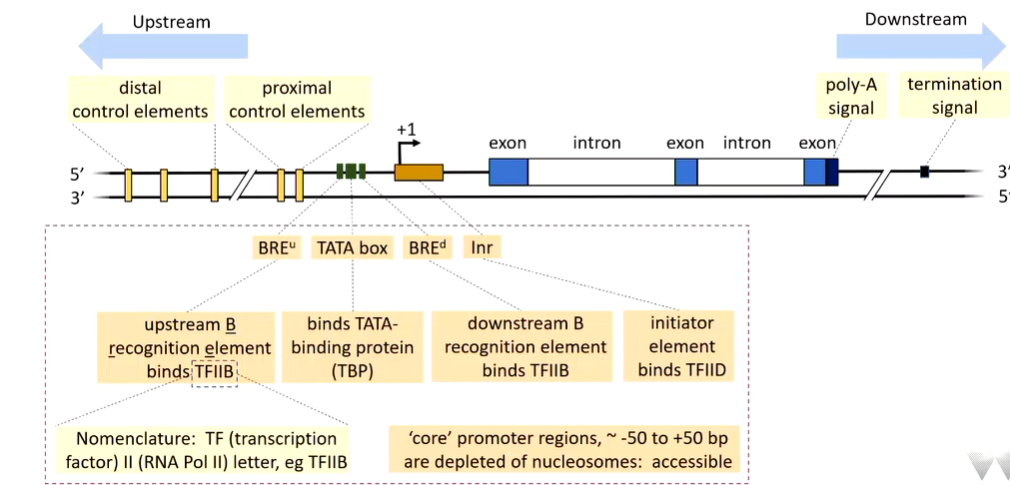
Pol II does not recognise this promoter directly. The TATA box is normally required. Upstream of this is the BRE which binds TFIIB (transcription factor rna pol II). There is also a downstream BREd. These flank the TATA box.
Distal control elements are regarded as enhancers although they do the same stuff as proximal ones. These govern the change in structure of the chromatin
Not all promoters look like this as they are variable.
LF104: 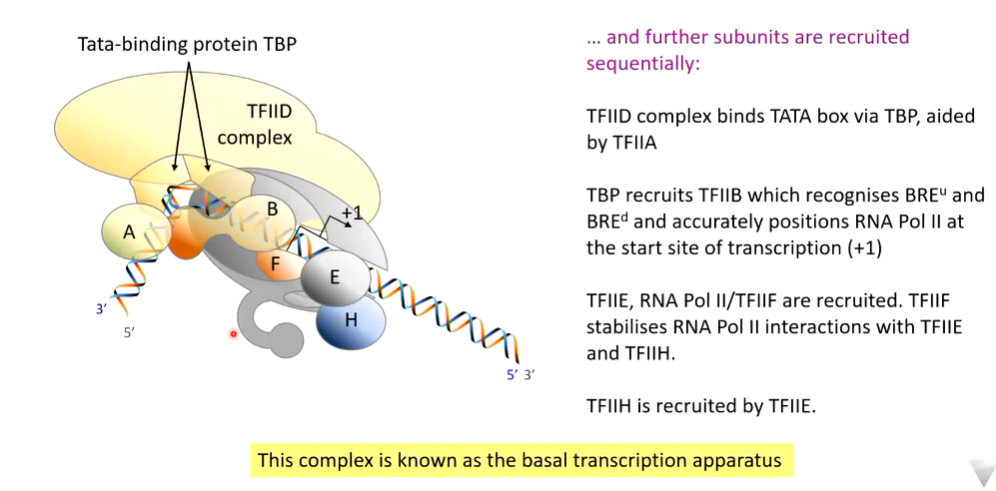
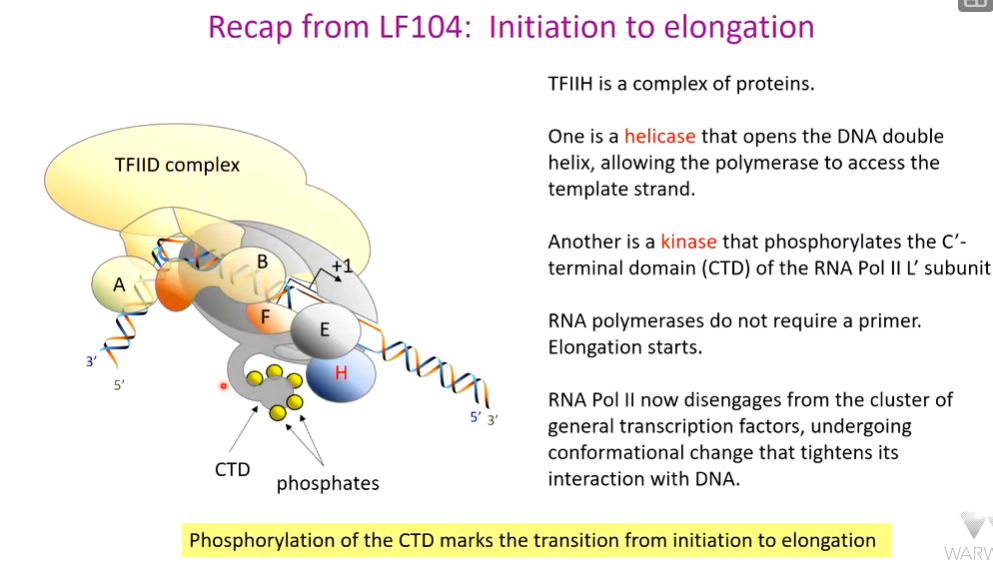 the TATA box is a consensus sequence and individual TATA boxes have different affinities for TBP - and so some are more efficient at stimulating transcription than others.
the TATA box is a consensus sequence and individual TATA boxes have different affinities for TBP - and so some are more efficient at stimulating transcription than others.
The G-less cassette is an artificial piece of DNA that lacks G residues. It is cloned downstream of a promoter. At one point there is a G residue from the original DNA. All the components for transcription are provided. The new RN copy is only made up of ACU means the RNA transcriptions stops when a C residue is reached as there is no G provided. If the uridine is radioactive, the RNA is radioactive and so can be quantified. Basically because no GTP is supplied, the RNA is truncated at the point at which the G should be inserted.
The sequence of a strong TATA box can be taken (major late adenovirus promotor - human viral promoter). If the three nucleotides are supplied with no G and transcription factors, there is no transcription as the TFIID complex has to be supplied. If this is supplied, there is strong expression of the G less cassette. If the end of the TATA box is mutated into a G, the human TFIID binds less strongly to the TATA box and there is reduced expression 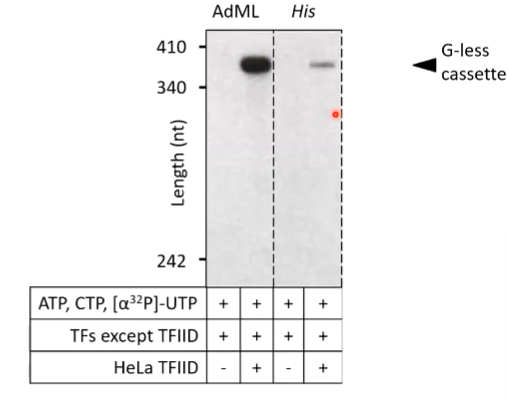
(His = yeast His TATA box). The ability to find the TATA box depends on the sequence of it - 1st level of transcriptional control
This can be extended into further studies - epstein barr virus productive cycle. Late genes have a distinct TAATT motif and early intermediate (IE) and early (E) have a TATA motif. This shows temporal control as there is control dependant on the time the gene is expressed. IE and E genes are differentially expressed so there are other control sequences
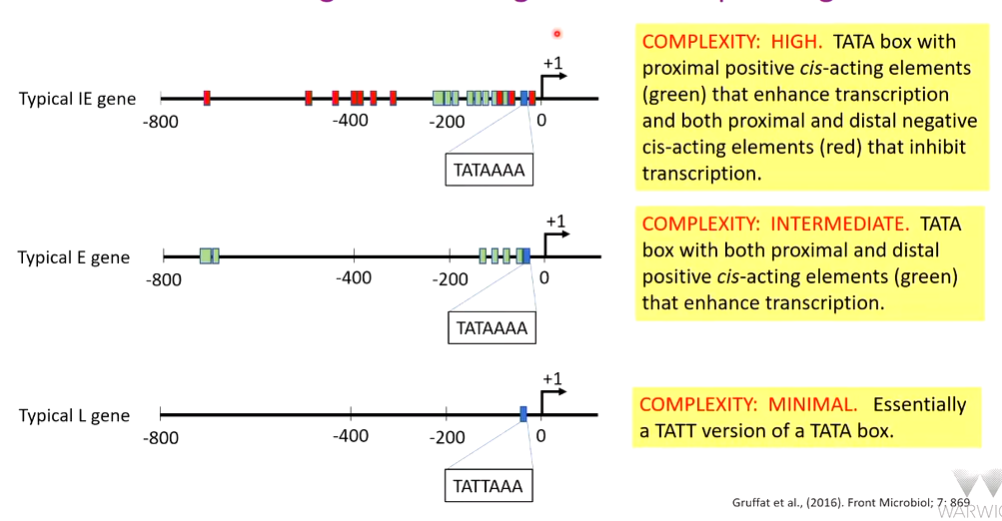
Additional binding and activation factors
These are activator/repressor proteins, mediator proteins and chromatin modifying proteins.
Eukaryotic RNA polymerases cannot access DNA selectively and so require additional factors. 
Homeodomains
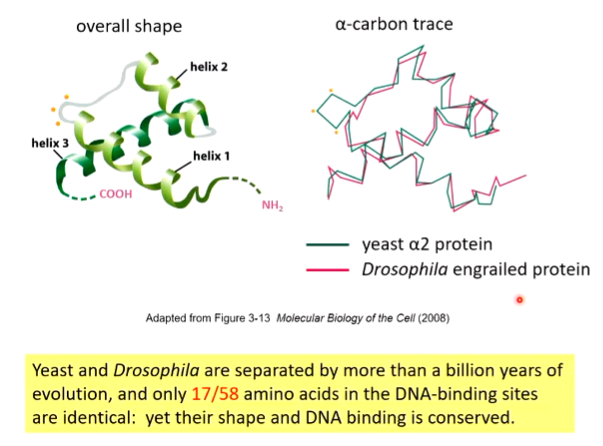
There is a conservation of shape as the long helix 3 sits in the major groove of DNA and makes specific contacts with specific base pairs - base specific interaction with DNA through Ser, Arg and Asn
Zinc fingers
characterised by two beta strands and and an alpha helix. The two beta sheets bind one zinc molecule and the two histidine residues also bind this. These coordinate the zinc molecule which stabilises the fold to interact with the major groove. 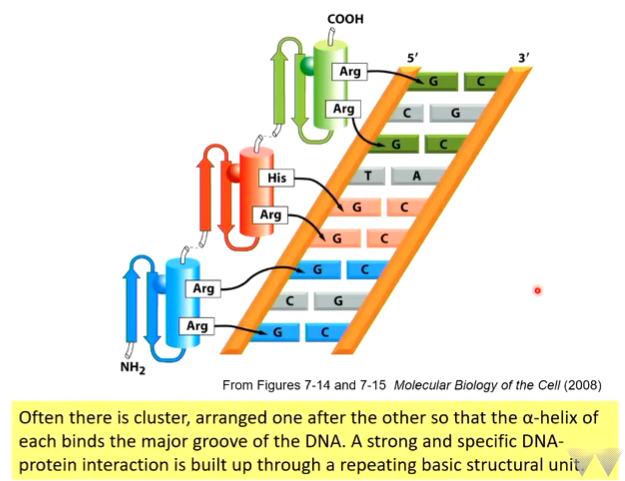
Specific contact with G residues. GC rich areas are prime binding site areas for this - base specificity
Leucine zipper
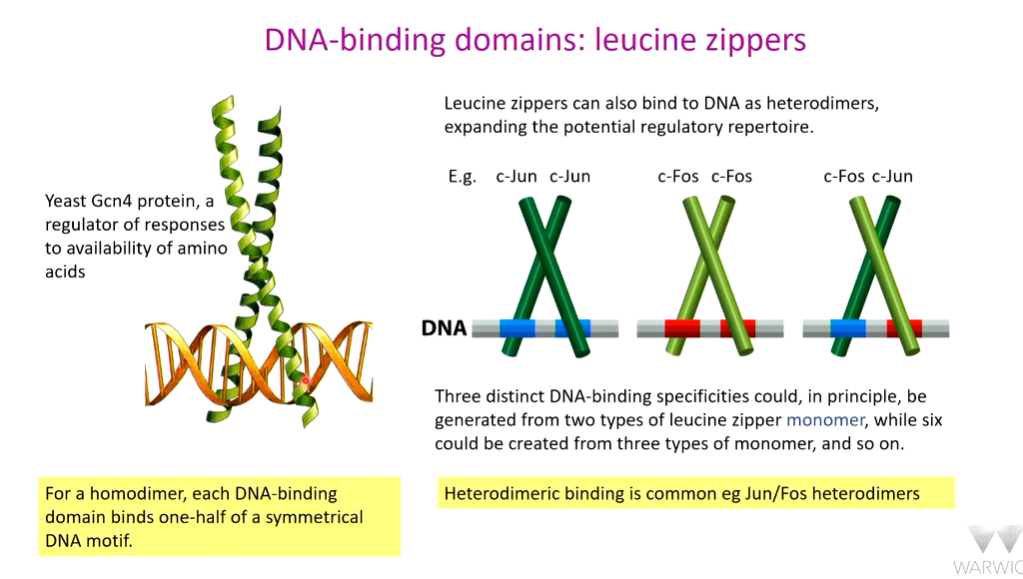
This allows tight and regulated expression of key genes in a tunable fashion. Signalling through Jun Fos pathway -target site for T cells. NFAT stimulate low level expression of IL-2. If all three pathways are activated, there is strong expression of IL-2. 
enhanceosome
There is tight regulation of DNA expression
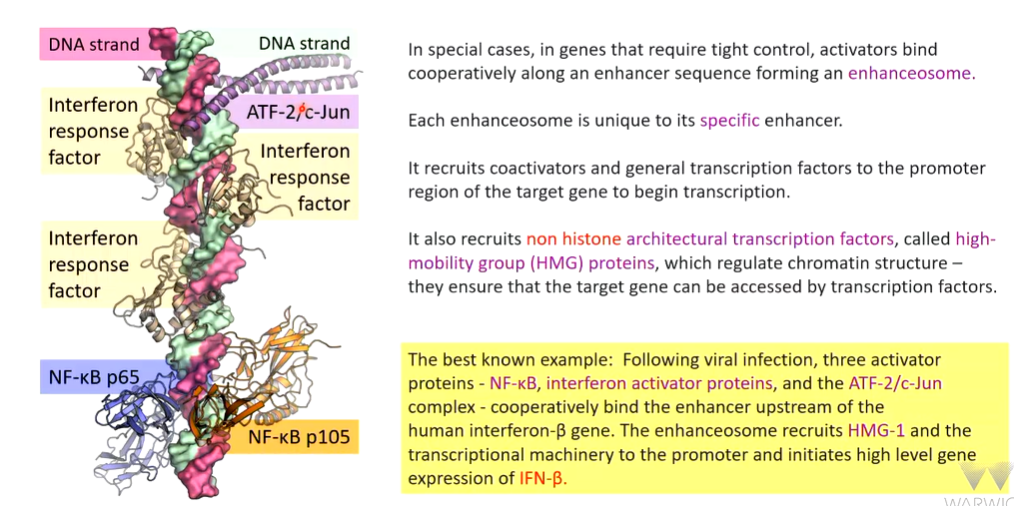
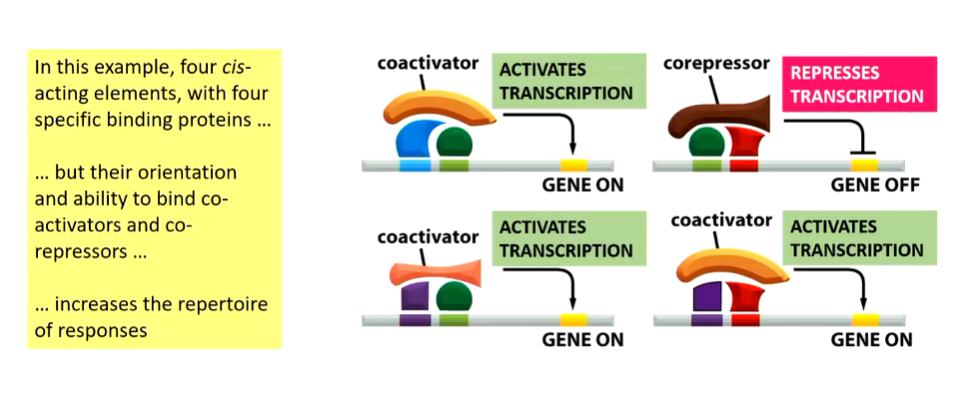
When this goes wrong the philadelphia chromosome occurs which is the gene translocation effect between Chr 9 and 22 that brings the ABL1 and BCR genes together. 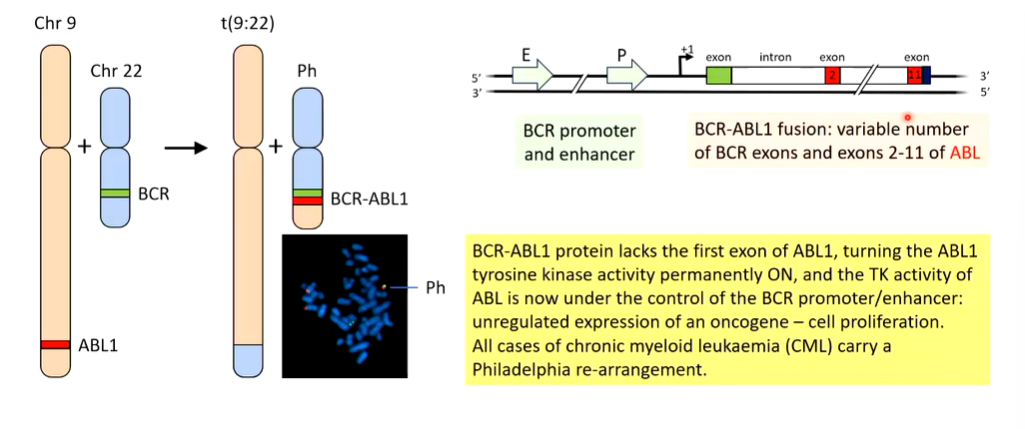
Burkitt’s lymphoma can also occur which is a translocation event between Chr 8 and 14: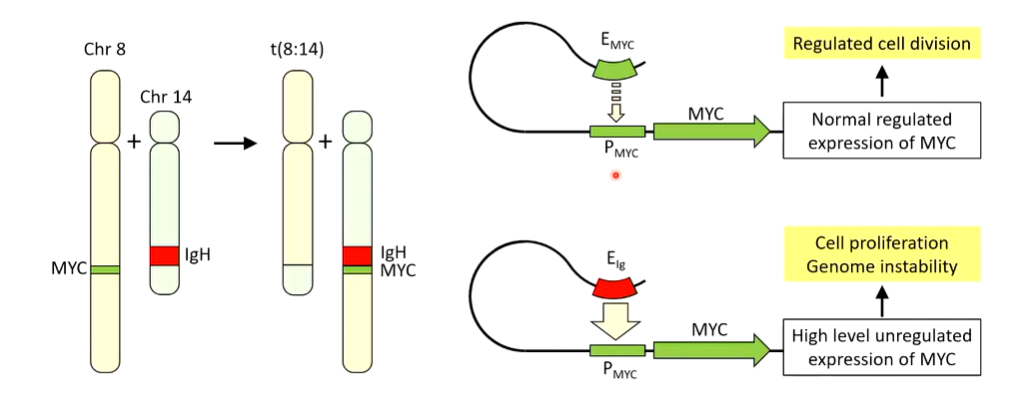
EXAM: 