Contração muscular
1/69
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced |
|---|
No study sessions yet.
70 Terms
células musculares
São células capazes de dar como resposta diversos tipos de estímulos, sejam eles químicos, elétricos ou mecânicos, sendo também capazes de converter a energia da hidrólise do ATP em energia mecânica, ou seja, num movimento de uma fibra sobre a outra, originando o movimento do órgão a que estão ligados.
Podemos definir 3 tipos de células musculares:
• Esqueléticas: constituem a maior parte da massa muscular do organismo e apresentam estrias transversais, estando ligadas a movimentos voluntários.
• Cardíacas: apresentam estrias e vão responder a uma estimulação rítmica a partir de células pacemaker do coração.
• Lisas: sem estrias, encontram-se na maior parte dos órgãos viscerais ocos, formando um sistema que tem contrações irregulares contínuas que não dependem da inervação.
Características do Músculo
O músculo apresenta um conjunto de propriedades e características básicas que são necessárias ao seu funcionamento.
• Excitabilidade: o músculo é capaz de responder a estímulos químicos, mecânicos (ex: de distensão) e elétricos.
• Condutividade: capazes de transmitir esses estímulos ao longo da sua fibra, permitindo depois o início da contração.
• Contractilidade: capazes de encurtar quando estimulados.
• Extensibilidade: capazes de aumentar o seu comprimento até determinado valor.
• Elasticidade: capazes de retornar ao seu comprimento inicial após a distensão.
Músculo Esquelético
É um músculo estriado de ação voluntária que se encontra ligado ao osso através de tendões, apresenta fibras musculares longas com bandas claras e escuras que se
parecem com estrias, e está sob controlo consciente.
Estas fibras individuais começam e terminam nos tendões, e estão dispostas em paralelo com as terminações dos mesmos, o que permite que a que força desenvolvida pelas unidades seja aditiva e, portanto, permite desenvolver uma capacidade de força muito maior.
Cada fibra é uma célula simples, multinucleada, longa e com formato cilíndrico, circundada pela membrana celular que aqui se designa sarcolema. Cada fibra é depois formada por um conjunto de miofibrilhas, invisíveis em filamentos individuais, constituídos pelas proteínas contrácteis.
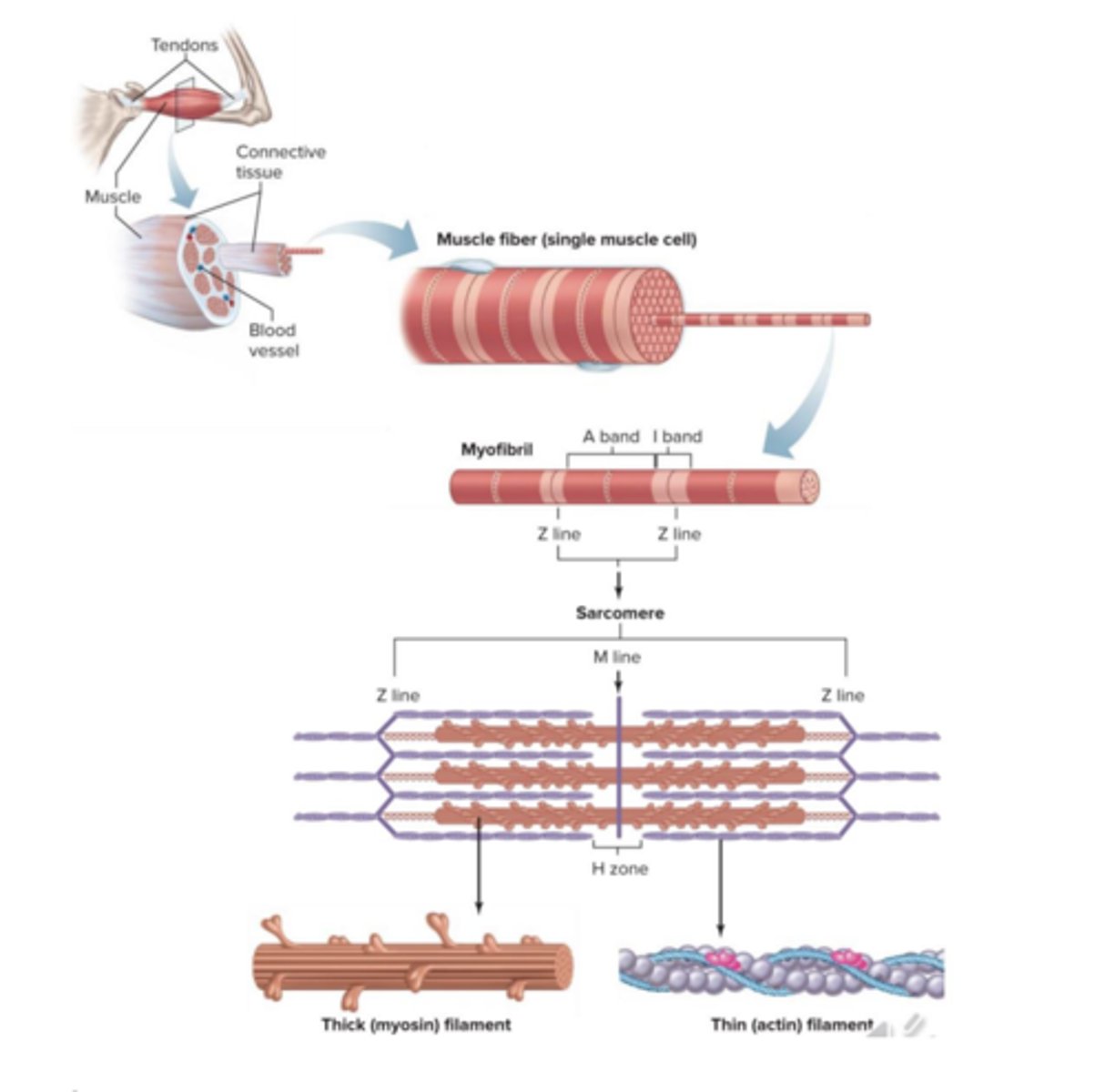
Fibras Musculares
As fibras musculares contêm múltiplos núcleos dentro da
membrana plasmática. As proteínas contrácteis são a miosina, actina, tropomiosina e a troponina.
O sarcolema destas células tem túbulos transversos que penetram a célula e que permitem a propagação do estímulo para o interior da mesma. Estes túbulos vão conseguir chegar mesmo ao interior da célula e isso vai ser muito importante na propagação dos estímulos que vão iniciar a contração.
O sarcoplasma está cheio de miofibrilas designadas miofilamentos. O retículo sarcoplasmático destas células contém uma grande quantidade de cálcio, que vai também ser muito importante no mecanismo contrátil.
Estrias
O músculo esquelético apresenta um padrão de estrias transversais. Assim, é possível definir um conjunto de bandas, dependendo da sua coloração ou da sua refração ser mais clara ou mais escura.
Podemos definir a banda I, mais clara, correspondendo à zona com os filamentos finos.
A banda A, mais escura, correspondendo à zona com os filamentos grossos e à sobreposição destes filamentos com os filamentos finos. No interior da banda A conseguimos definir, quando o músculo está em relaxamento, uma banda mais clara, banda H, (zona em que apenas existem filamentos grossos, não existindo sobreposição entre filamentos finos e filamentos grossos).
Podemos definir também uma linha Z, mais escura, correspondendo a uma rede de proteínas de ancoragem onde se vão ligar os filamentos finos.
Por fim, podemos dizer que a área correspondendo entre linhas Z, é definida como um sarcómero.
O arranjo das fibras contráteis está principalmente assente em dois principais tipos de filamentos:
os filamentos grossos de miosina e os filamentos finos de actina.
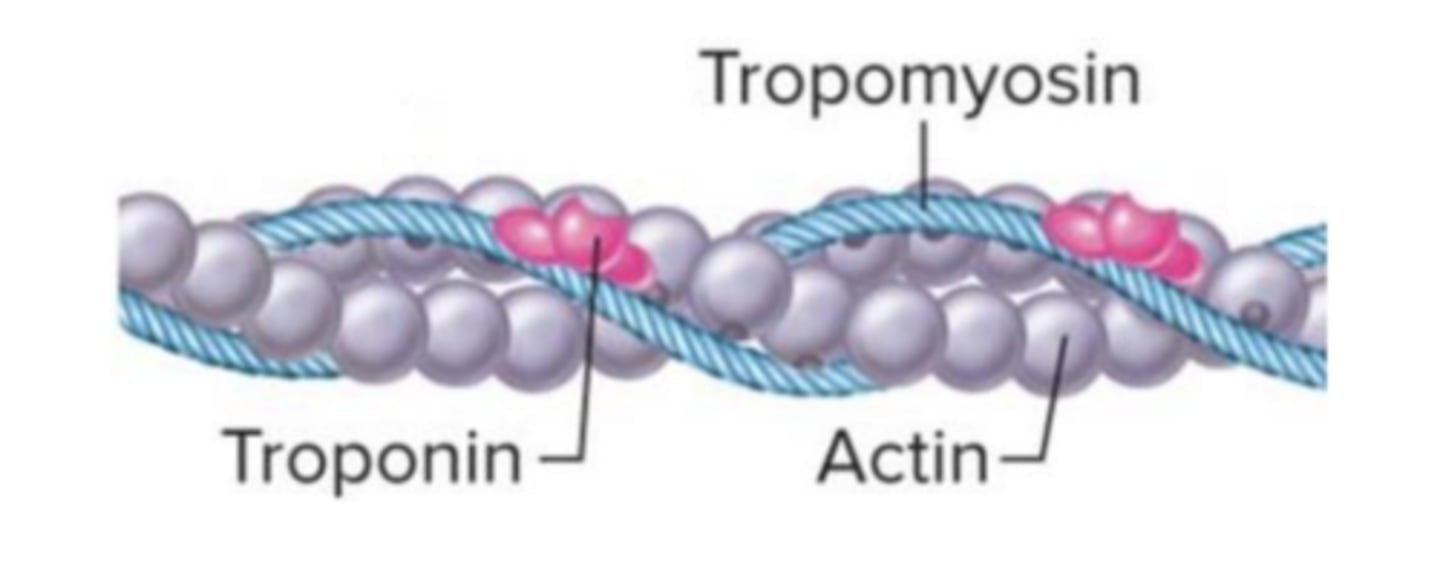
Filamentos Finos
São formados por actina, tropomiosina e troponina.
Estes filamentos de actina apresentem uma organização em dupla hélice longa, nas quais assentam as moléculas de tropomiosina, que são também eles filamentos longos
localizados nos espaços entre as duas cadeias de actina; cada filamento destes contém cerca de 300 a 400 moléculas de actina e 40 a 60 moléculas de tropomiosina.
As moléculas de troponina são unidades globulares pequenas que se encontram em intervalos ao longo das moléculas de tropomiosina, sendo formadas por 3 subunidades:
• Troponina T: faz a ligação à tropomiosina;
• Troponina I: inibe a interação da miosina com a actina;
• Troponina C: contém os locais de ligação do cálcio.
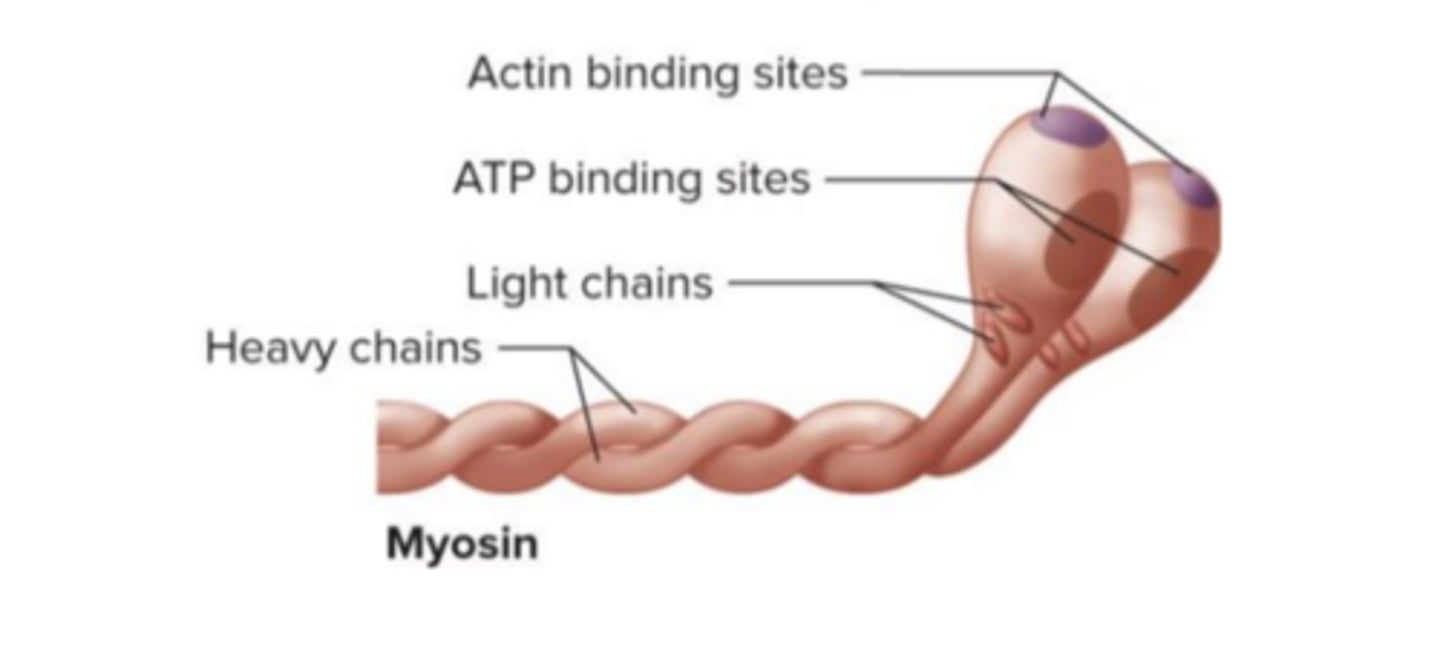
Filamentos grossos
Os filamentos grossos são formados por miosina do tipo II e têm um diâmetro cerca de duas vezes superior ao dos filamentos finos.
Estes filamentos de miosina têm duas cabeças globulares e uma cauda longa; são essas cabeças que formam as pontes cruzadas com a actina. A cabeça de miosina forma ligações cruzadas com a actina e, para além do local de ligação à actina, tem um local que hidrolisa o ATP, comportando-se como enzima. Cada filamento destes contém cerca de 200 a 500 moléculas de miosina e forma uma estrutura semelhante a um taco de golfe.
Filamentos Elásticos e Ancoragem
A estabilização e estrutura do sarcómero é dada pela existência de discos Z formados por uma proteína de ancoragem designada por alfa actinina.
A capacidade de elasticidade existente no músculo esquelético é dada por uma proteína chamada titina (anteriormente designava-se por conectina), que liga a miosina ao disco Z.
Esta é a maior proteína existente na natureza, e permite manter alinhados os filamentos finos e grossos, resistir à distensão e manter a banda A no centro do sarcómero.
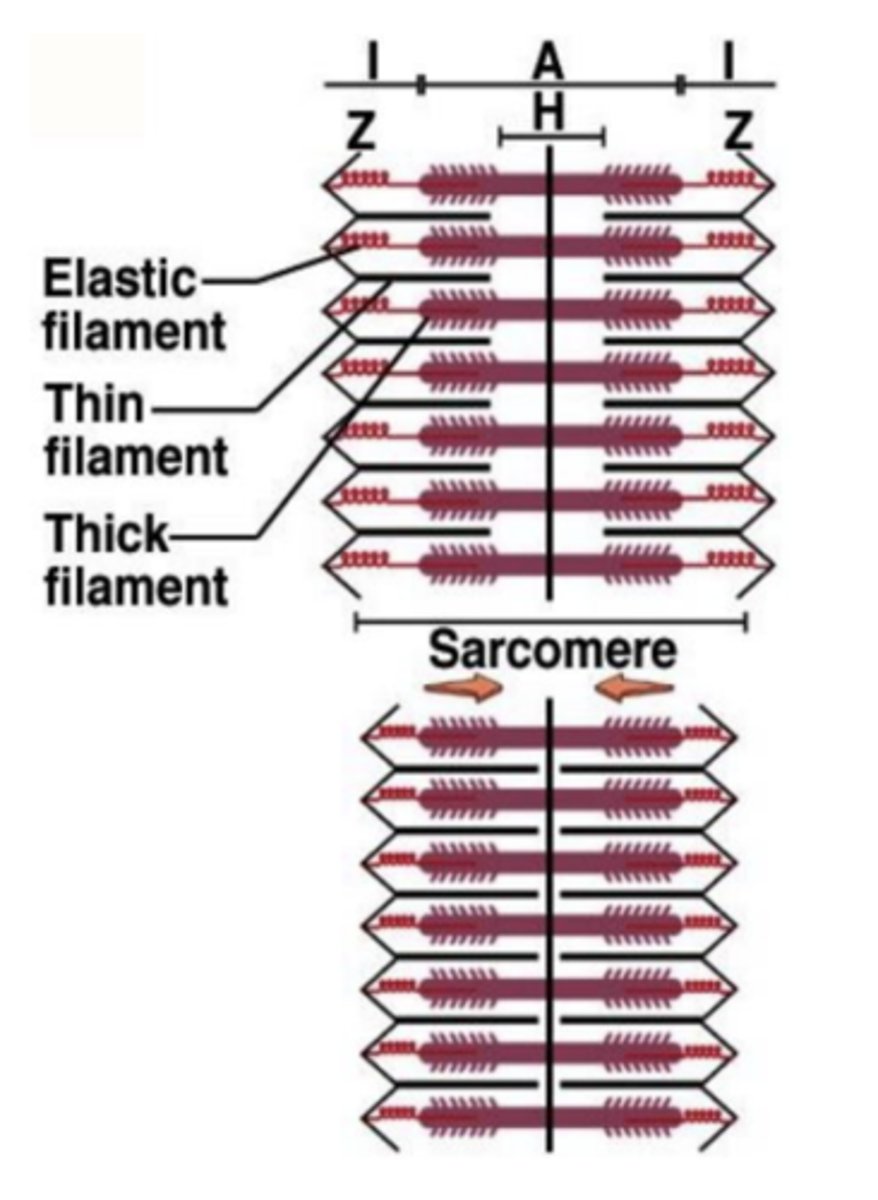
Relaxamento vs Encurtamento
Numa contração muscular, a força desenvolvida pelo músculo é devida ao encurtamento do comprimento do sarcómero, com o desaparecimento da banda H.
Junção Neuromuscular
A ligação entre o nervo e o músculo é feita pela junção neuromuscular. Neste tipo particular de sinapse, os potenciais de ação que estimulam o músculo esquelético são gerados em neurónios motores que têm o seu corpo celular localizado no tronco cerebral ou na espiral medula. Estes neurónios são mielinizados, e por isso a velocidade de condução é elevada.
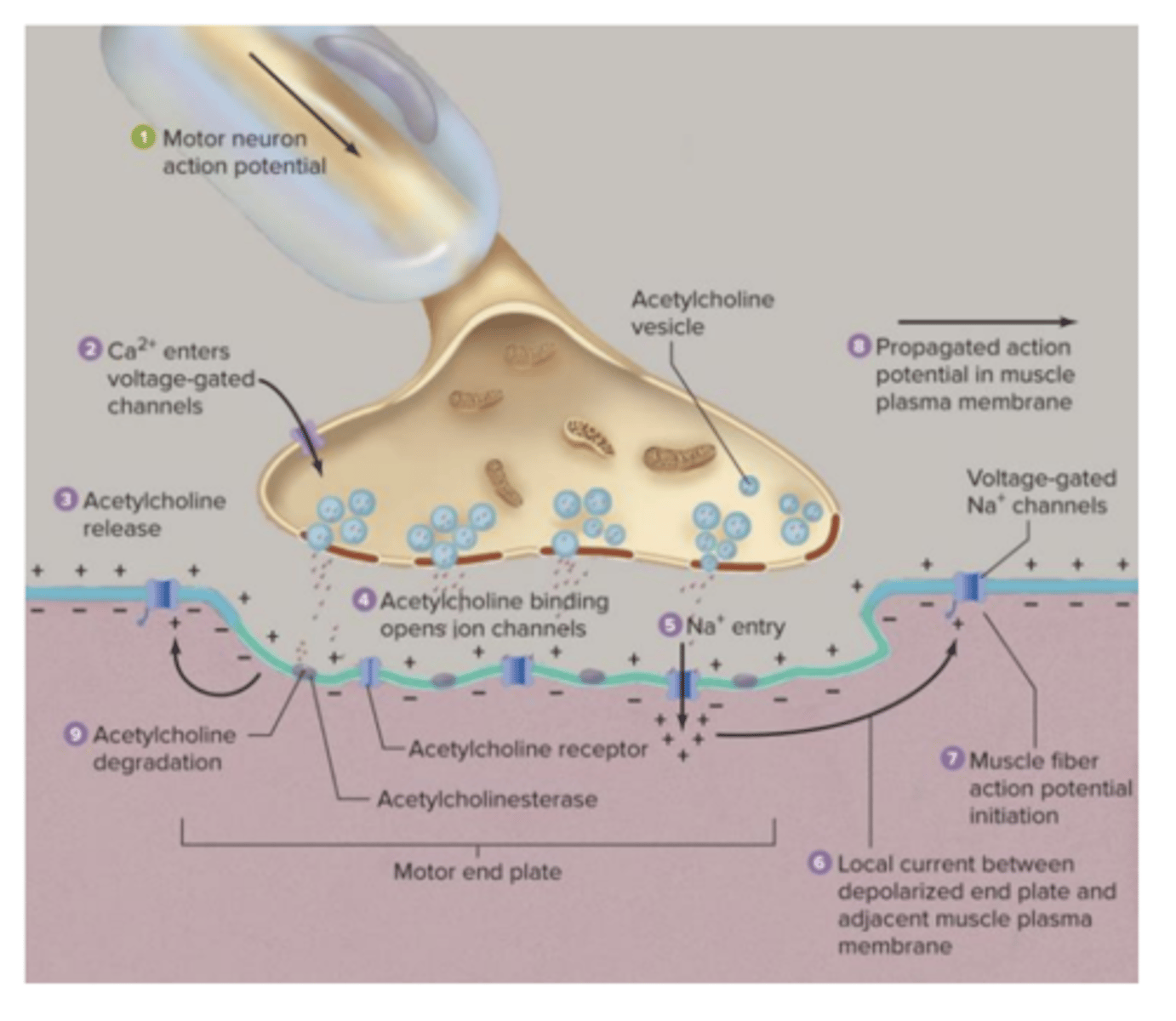
Junção Neuromuscular - esquema

potencial de repouso
do músculo esquelético é cerca de -90mV; o potencial de ação durará entre 2 a 4 ms, e este será conduzido ao longo da fibra muscular com uma velocidade de cerca de 5 m/s.
Este músculo irá apresentar um período refratário absoluto entre 1 a 3 ms.
potencial de ação isolado
provoca uma contração, seguida de um relaxamento; essa sequência de acontecimentos é designada de abalo muscular. Este abalo começa cerca de 2 ms após o início da despolarização, e a sua duração vai depender do tipo de fibras.
• Nas fibras rápidas, o abalo muscular tem uma duração de cerca de 7.5ms.
• Nas fibras musculares lentas, os abalos musculares rondam a ordem dos 100ms.
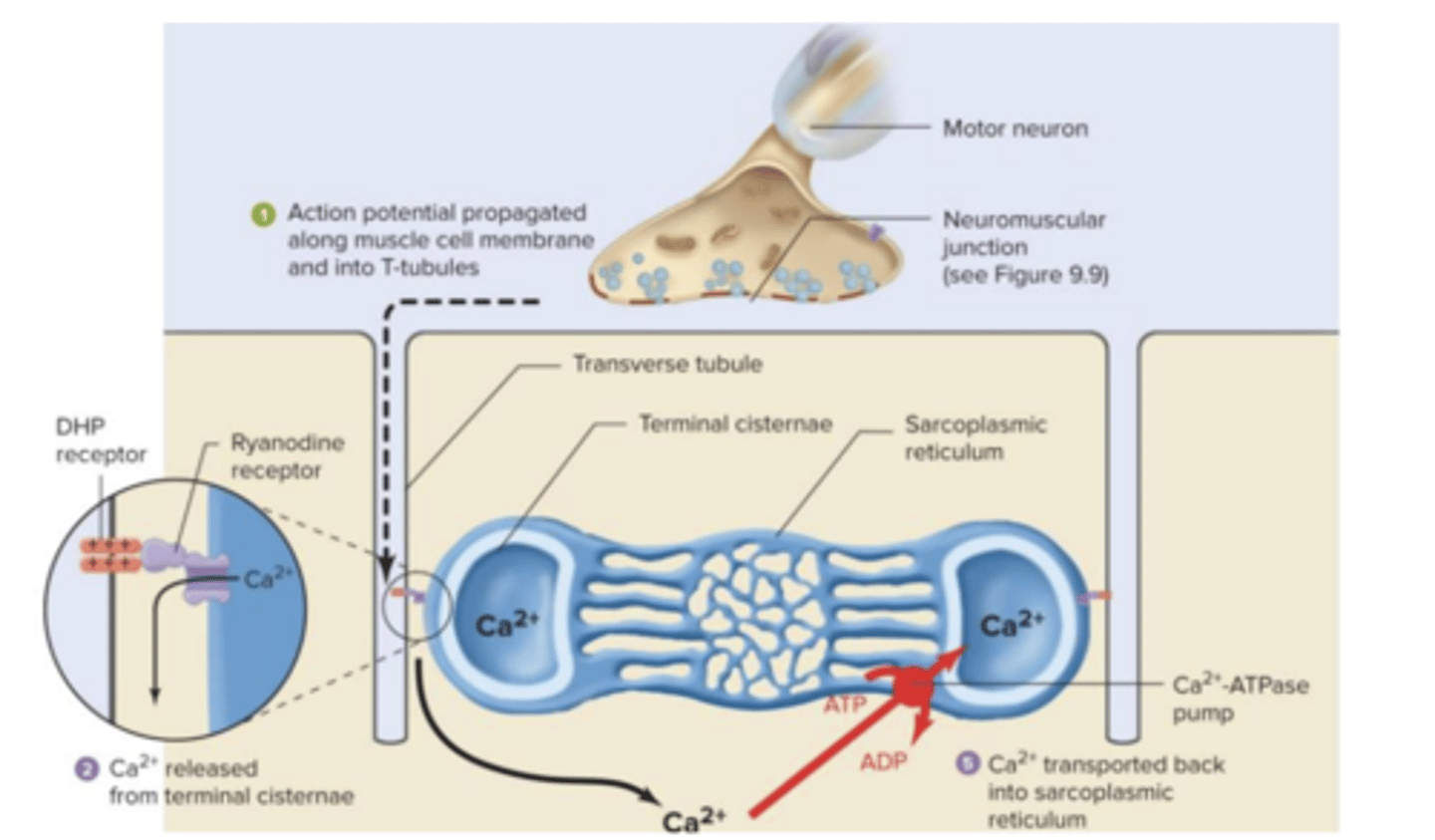
Acoplamento Excitação-Contração
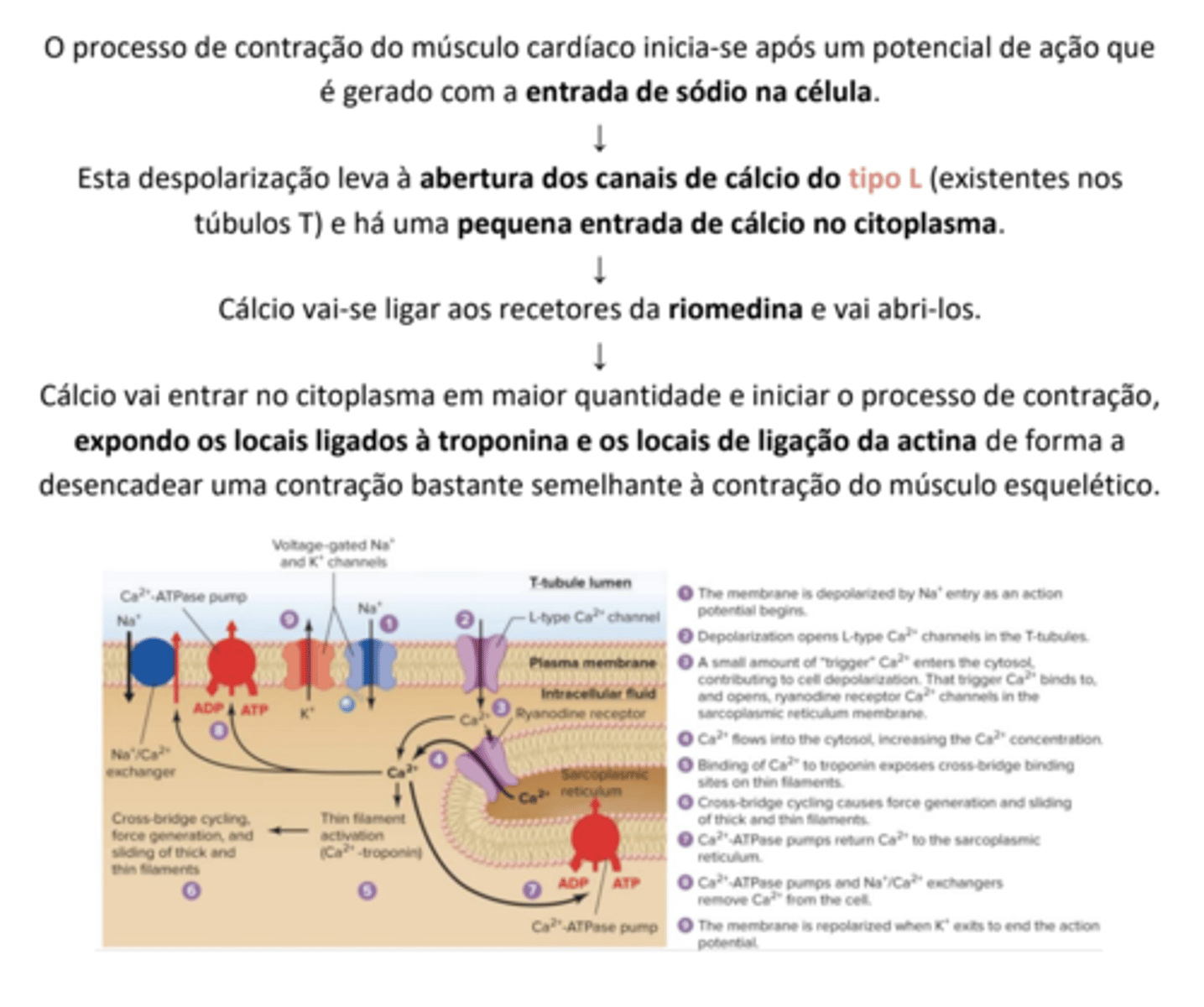
O processo pelo qual a despolarização da fibra muscular inicia o processo de contração designa-se por acoplamento excitação-contração.
Após o início do potencial de ação no músculo, este é propagado e chega aos túbulos T onde penetra para o interior da fibra.
Um mecanismo especializado liga os potencias de ação existentes nos túbulos T à libertação de cálcio pelo retículo sarcoplasmático - os túbulos T estão intimamente ligados ao retículo sarcoplasmático através de um sensor de voltagem chamado diidropiridina, que por sua vez se encontra ligado ao recetor da rianodina.
Quando o potencial de ação chega ao recetor da diidropiridina, os resíduos de aminácidos carregados deste recetor induzem uma alteração conformacional do recetor da rianodina, que por sua vez atua como um canal de cálcio que abre e provoca a libertação do cálcio das cisternas terminais, permitindo a saída de cálcio livremente para o citoplasma, e iniciando-se depois a contração através do ciclo das pontes
cruzadas.
Para que se inicie a ligação da miosina à actina, é necessário que os locais de ligação se encontrem desimpedidos. O cálcio libertado do retículo sarcoplasmático liga-se à troponina C, provocando um enfraquecimento da ligação da troponina I à
tropomiosina e permitindo que ela se movimente lateralmente. Este tipo de movimento descobre os locais de ligação para as cabeças de miosina e a remoção do cálcio do meio reverte este processo.
São descobertos cerca de 7 locais de ligação da miosina por cada molécula de troponina C ligada ao cálcio.
vias piramidais
1º neuronio origina-se no giro pre-frontal, passa pela via cortico-espinhal e cortico-bulbar, cruzam para a posição contraria decussação, 75%
cortico espinhal, controla musculos abaixo do pescoço
cirtico bulbar, controla pescolo e face
2º neuronio pode começar no tronco ou na medula espinhal, que termina nos musculos esqueleticos alfa motores
vias extrapiramidais
potencias veem debaixo do cerebro
controlo priamidal
nao oassa nas piram idades do bulbo
são4
intensificam ou deseintensificam o controlo voluntario
se for mt intensificado o individuo perde o controlo
unidade motora
conj do neur (fibra nervosa) alfa motor, controla fibras musculares
fibras esqueleticas sao finas
qt menores as unidades de um musculo mais preciso é o mov realizado
musc postorais - und motoras grandes
recrutamento de unidades motoras
apenas aquelas q são necessárias
começando por recrutar as mais pequenas e s epor feedback vai fazer. mais força recrutando unidades maiores
contração
os pontos onde se fixa aproxiamam.se, inserções aproximam-se (quase todos em ossos)
Em repouso, a miosina não consegue ligar-se à actina devido à reduzida concentração de cálcio.
No entanto, as cabeças de miosina já se encontram energizadas, devido à hidrólise do ATP em ADP e fosfato inorgânico.
Quando o cálcio entra no citoplasma, a miosina liga-se à actina e o ciclo inicia-se.
A ligação da miosina energizada à actina provoca o movimento da cabeça e consequente deslizamento da actina sobre a miosina; a energia armazenada na cabeça de miosina
antes da ligação entre os dois filamentos é posteriormente libertada durante esta ligação, provocando assim o movimento.
Durante o movimento das pontes cruzadas, a miosina está fortemente ligada à actina, e só com a quebra desta ligação se consegue re-energizar a cabeça de miosina e repetir o ciclo.
Este processo requer a ligação de uma molécula de ATP à miosina para quebrar esta ligação; a molécula de ATP vai funcionar como um regulador alostérico e não como uma fonte de energia, uma vez que a ligação desta molécula diminui a afinidade da miosina para a actina e assim permitir a quebra da ligação.
Após a dissociação da ligação entre a actina e a miosina, o ATP é hidrolisado, energizando a cabeça de miosina e iniciando o ciclo novamente.
O ATP tem assim dois papéis importantes: por um lado é a fonte de energia para o movimento em si, e por outro lado é o regulador alostérico para o desacoplamento entre a actina e a miosina.
musculo relaxado
actina coberta com tropomiosina
troponina decide pegar na tropomiosina e possibilita a lig cruzada/ponte entre a actina e a cabeça da miosina
O processo de relaxamento após a paragem do estímulo ocorre quando deixa de ser libertada acetilcolina para a fenda da junção neuromuscular e, consequentemente, as acetilcolinesterases que já existem nessa fenda degradam a acetilcolina, até que ela desapareça dessa fenda ou que reduza substancialmente a sua concentração.
Desta forma, o número de neurotransmissores é muito reduzido e deixa de iniciar um potencial de ação no músculo, uma vez que não se atinge o limiar de excitação.
Uma vez que não existe potencial de ação, o cálcio deixa de ser libertado do retículo sarcoplasmático e é bombeado novamente para as zonas das cisternas terminais através de bombas ativas de cálcio ATPases, que o vão armazenar nessa zona, preparando-o para uma nova contração.
A perda de cálcio do citoplasma leva à remoção do cálcio da troponina C, o que provoca a alteração da conformação da tropomiosina, bloqueando novamente os locais de
ligação da actina. Isto leva a um completo relaxamento, enquanto não existir novamente mais cálcio no meio.
placa terminal
terminação do neuro-alfa motor, recetores colinergicos nicotínicos pois aceitam nicotina e acetilcolina
ponbtecial de acao
entre esq e motor difere pontecial de repouse esq nao tem hiperbolização
tubulos T
musc excitado pela acetilcolina, depois propagação ´e feita horizontaletne e perpendicular ao eixo maior em proufundidade. invaginaçãp da membrana plasmática
nas cel esq o orgnaleo q liberta calcio, reticulo end está mt proximo dos tubulos t estanto proximo da membrana plamsmatica
transformação informação elétrica para mecânica
dá-se a nivel do túbulo t a partir de canas de voltagem de cálcio q n transportam calcio
faz -se na ligação entre os dois canais de calcio
reanodicnico possibliza de calcio passe a favor do gradiente de concentração, ou seja sai do retículo endoplasmático
ciclo pontes
dita se está a contrair (actina exposta e miosina tem q ter afinidade atraves da relaçãocom o atp, hidrolisado em adp e fofofato inorganici , cada cabeça empurra actina para o centro do sarcomero depois soltam-se) ou relaxar (quebra-se lig atraves do s+atp com cabeça de miosina e delvove-se clacio ao reticulo através de transportadores ativos, SERCA transporta atp asseguram mov de calcio)
abalo muscukar
contracao de uma unica celulas muscular
fase de latencia - ordenar saida de calcio do reticulo onde ocorre potencial de acao
fase de contração
fase de relaxamento - calcio de volta ao reticulo
somação temporal de abalos musculasres
músculo faz mais força gerando mais ponteciais de ação, concentrando mais calcio
tetania
força máxima assegurada pelo musculo
tetania nao fundida - fase incial
tetanio fundida - cada abalo fundio-se com o seguinte
fadiga muscular
deixa de contrair apesar de estar a e ser estimulada
fadiga periferica
acumulação toxicos acidos latico ou redução atp e falta de glicogenio
fadiga central
nao estar motivado
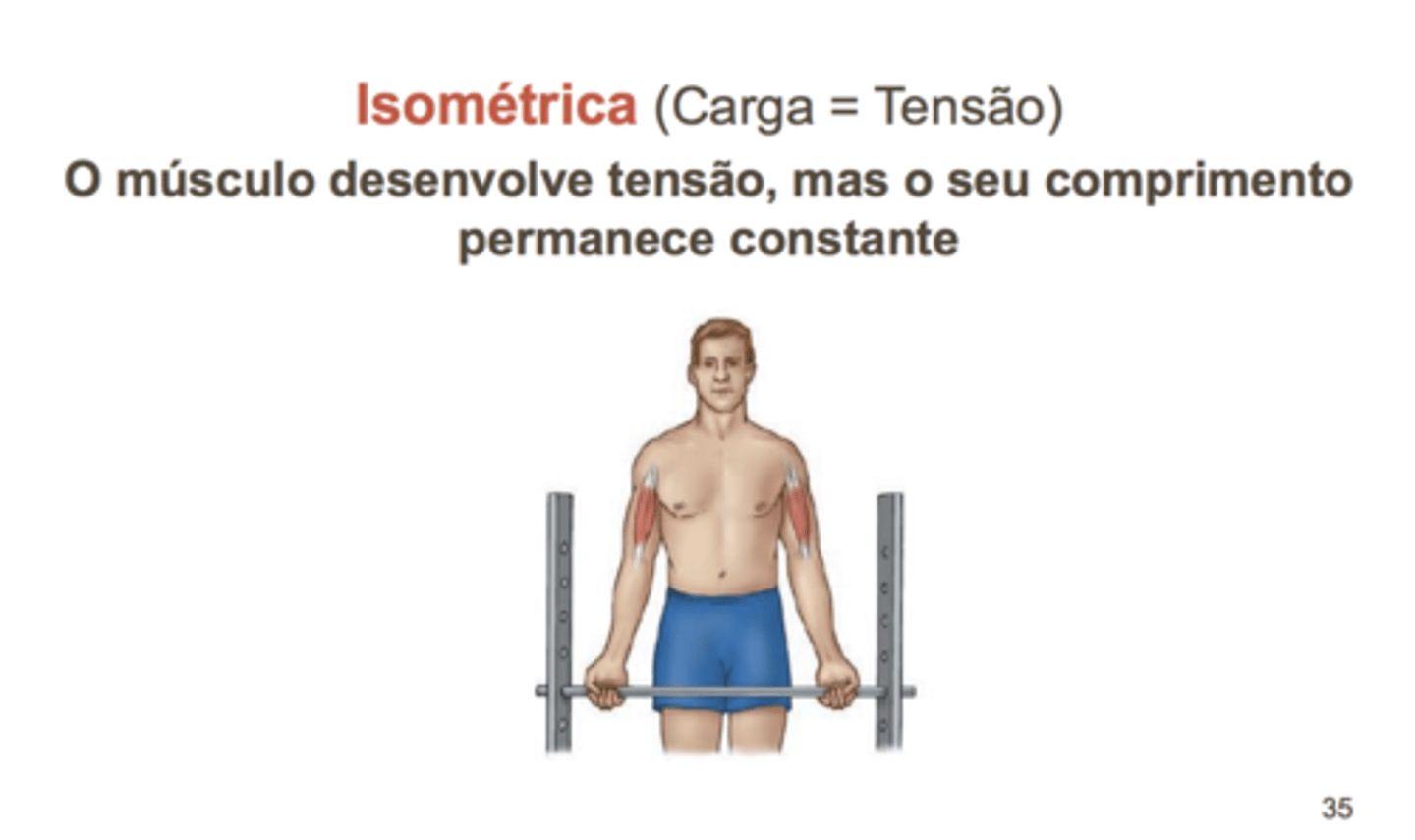
Tipos de Contração
A contração muscular envolve o encurtamento dos elementos contráteis; no entanto, como o músculo apresenta elementos elásticos e viscosos ligados em série com as fibras contráteis, é possível desenvolver uma contração sem redução do comprimento do músculo.
Neste caso, esta contração designa-se por contração isométrica, não havendo encurtamento nem gerando trabalho.
A contração que se realiza contra uma carga constante, com aproximação de extremidades do músculo, designa-se por contração isotónica, sendo uma contração capaz de gerar trabalho.
Propriedades da Contração Muscular
Após um potencial de ação, existe um período de alguns milissegundos até que a tensão das fibras musculares comece a aumentar - período de latência. Durante este período,
o processo de acoplamento excitação-contração está a decorrer.
Comparando os abalos musculares isométricos e isotónicos numa mesma fibra muscular, verifica-se que o período de latência de uma contração isotónica é maior do
que nunca contração isométrica; no entanto, a duração do movimento mecânico é inferior do que numa contração isométrica.
Isto deve-se ao facto de, numa contração isométrica, a tensão se iniciar assim que se ligam as primeiras pontes cruzadas; logo, o período de latência deve-se apenas ao atraso
do acoplamento excitação-contração.
Já numa contração isotónica, o período de latência inclui o tempo de início do acoplamento excitação-contração, e existe também um tempo extra para que a fibra muscular acumule pontes cruzadas suficientes para ter força suficiente para vencer a carga que está a ser movimentada; é por isto que o período de latência acaba por ser maior.
Isométrica
Isotónica
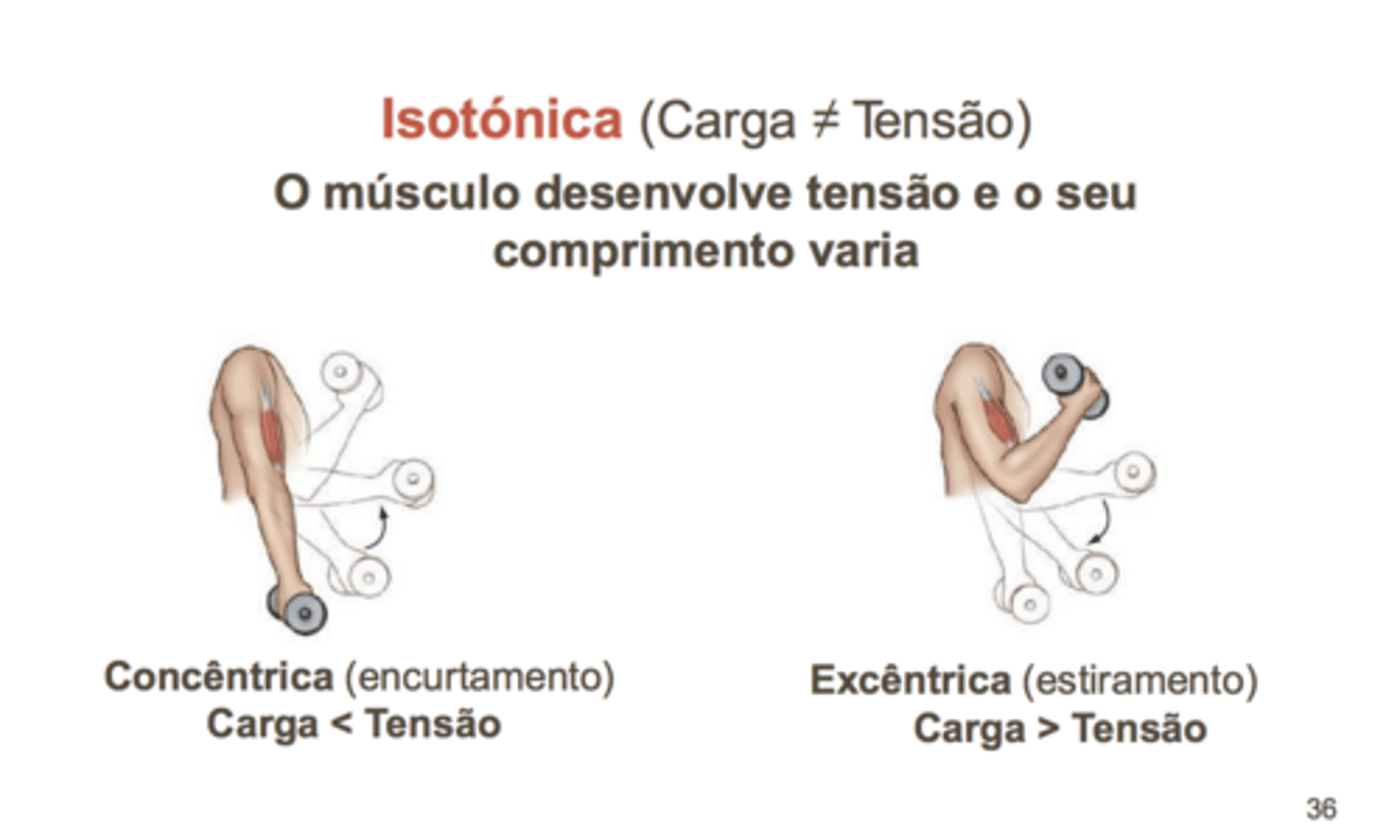
características de uma contração isotónica dependem da carga; a cargas
altas:
• O período de latência é alto;
• A velocidade de encurtamento é mais lenta;
• A duração do abalo muscular é também reduzida;
• A distância de encurtamento é inferior.
Para cargas mais altas, o músculo não começa a encurtar até que a carga seja vencida; assim, existe uma contração isométrica inicial antes de se iniciar o período de contração isotónica - um período de tempo em que o músculo está a acumular tensão, mas que é inferior à carga a deslocar.
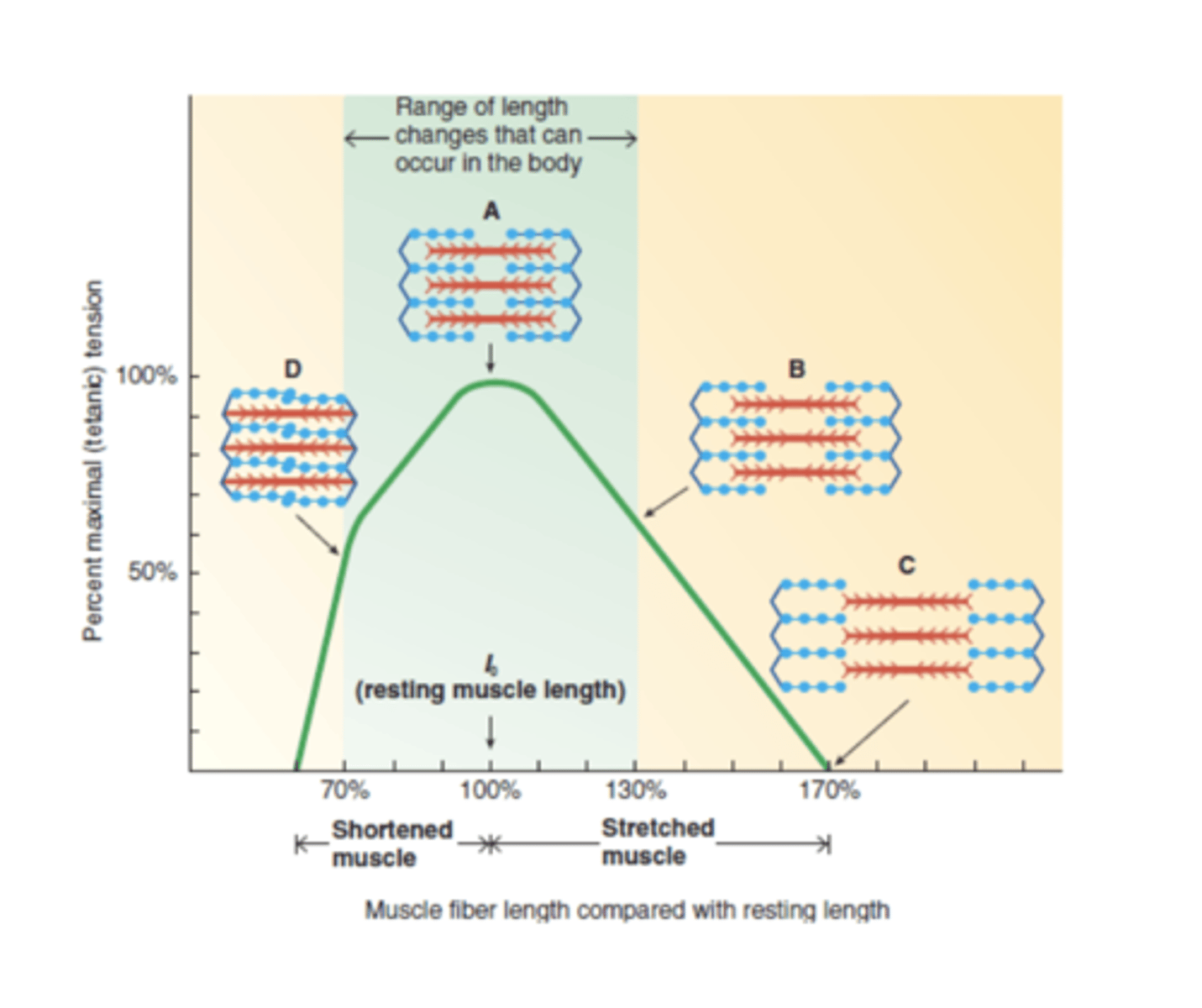
Relação comprimento - tensão
comprimento ótimo a verde
à esq do comp otimo
pontes cruzadas diminuem quando está demasiado comprimidos
à dir do comp otimo
finos e espessos nao se encontram bem e numero de pontes cruzadas devido à grande tração
tensão relação direta com nº de pontes cruzadas
Efeito da Estimulação sobre a Contração
Como um potencial de ação dura 2 a 4 ms mas o abalo muscular pode durar até 100 ms, é possível que se inicie um novo potencial de ação durante o período de atividade
mecânica. Assim, se dois estímulos sucessivos forem iniciados suficientemente afastados no tempo, o músculo tem tempo de desenvolver um ciclo de contração e relaxamento e os dois estímulos iniciarão contrações exatamente iguais.
Se o segundo estímulo for iniciado ainda quando o músculo não teve tempo de relaxar completamente, inicia-se uma nova contração, com um pico superior ao da contração anterior; diz-se que há somação.
A existência de somação depende da estimulação e da frequência de estimulação.
• Até cerca de 10 estímulos por segundo, a contração muscular ocorre sempre com relaxamento.
• Entre 10 a 20 estímulos por segundo, cada contração desenvolve mais tensão devido à não remoção completa do cálcio do citoplasma; há uma contração que ainda se mantém.
• Entre 20 a 40 estímulos por segundo, cada estímulo chega antes da contração anterior terminar, entrando antes do tétano incompleto.
• Após 40 a 50 estímulos por segundo, não há tempo para existir um relaxamento longo, e portanto as contrações fundem-se numa contração prolongada designada de tétano prefeito.
Relaxação Tensão-Comprimento e Tónus
A tensão desenvolvida pela fibra muscular depende do comprimento do músculo antes de ser estimulado.
Se tivermos um sarcómero pequeno (quando os filamentos grossos estão muito perto dos discos Z), o início da contração desenvolve-se com uma tensão mais baixa porque as fibras não podem encurtar tanto, uma vez haver uma grande sobreposição.
Quando o sarcómero é muito grande (grande afastamento entre os filamentos finos e grossos e não há uma completa sobreposição entre as duas fibras), vão-se desenvolver
poucas pontes cruzadas ligadas à actina, desenvolvendo menor tensão.
Rigor Mortis
A importância de cálcio e ATP para a contração muscular é bem visível na situação de rigor mortis. Este processo inicia-se 3 a 4 horas após a morte, tem um pico às 12 horas, e diminui nas 48 horas seguintes.
Explica-se pela deterioração do retículo sarcoplasmático, que se inicia imediatamente após a morte, e que leva à libertação maciça do cálcio para o citoplasma das fibras musculares, provocando a disponibilidade maciça dos locais de ligação à actina.
Como ainda existe ATP no meio, ele vai ser utilizado para energizar as cabeças de miosina e provocar uma contração; no entanto, devido à quantidade de cálcio libertado, o número de ligações é grande, e por isso é necessário um grande número de ATP para iniciar o relaxamento. C
omo o ATP foi gasto na primeira fase e as células já não conseguem produzir ATP, o que vai acontecer é que as fibras vão manter-se ligadas até que a degradação dos miofilamentos se inicie, o que ocorre normalmente após 12 horas e dura até às 48 horas seguintes.
Energia
A mioglobina fornece oxigénio para uma respiração aeróbia ilimitada. As vias que se iniciam após o metabolismo anaeróbio permitem o aporte de ATP e provêm da
transferência de um grupo fosfato inorgânico de outras moléculas, como acontece no
sistema de fosfagénio, no sistema de mioquinase e na creatinina quinase, que capta
grupos fosfato inorgânico da creatinina fosfato e os disponibiliza ao ADP para formar
ATP.
Fontes de ATP
Durante o exercício, os vasos sanguíneos dilatam para fornecer oxigénio ao músculo.
Quando o esforço muscular é muito grande, a nova síntese aeróbia de reservas energéticas não permite acompanhar a sua utilização; nestas condições, utiliza-se
inicialmente a fosforilcreatina.
Parte da síntese de ATP é feita utilizando a energia libertada pela decomposição anaeróbia da lactose em lactato. A utilização desta via é, no entanto, limitada, pois mesmo com a passagem de lactato para a corrente sanguínea, existe uma quantidade que fica no músculo, ultrapassando a capacidade de tamponamento do tecido e levando a uma consequente diminuição do pH, acabando por inibir as enzimas. Esta via anaeróbia permite realizar grandes esforços em curto espaço de tempo (ex: corrida de 100 metros).
A partir desse tempo, tem de se iniciar outras vias metabólicas para produzir ATP e manter a contração do músculo.
Músculo Cardíaco
O músculo cardíaco é específico do coração.
• É composto por células mais pequenas e finas, juntas por discos intercalares;
• Possuem junções gap que permitem a passagem do potencial de ação de umas células para as outras de forma direta;
• Têm um retículo sarcoplasmático menos desenvolvido;
• Têm túbulos T mais largos que admitem cálcio do fluido extracelular;
• Têm células pacemaker que iniciam a contração;
• Utilizam a respiração aeróbica, uma vez que têm mitocôndrias maiores.
Músculo Liso
• É composto por células fusiformes com um núcleo;
• Não apresenta estrias, discos Z ou sarcómeros;
• Não tem túbulos T;
• Cálcio provém exclusivamente do exterior;
• O controlo nervoso é autonómico e, portanto, é um músculo de controlo Involuntário.
- SEM TROPONINA
- ESTURAS ANÁLOGAS subsitiuem linhas Z

Estimulação1
musc liso comandado por sist autonomo
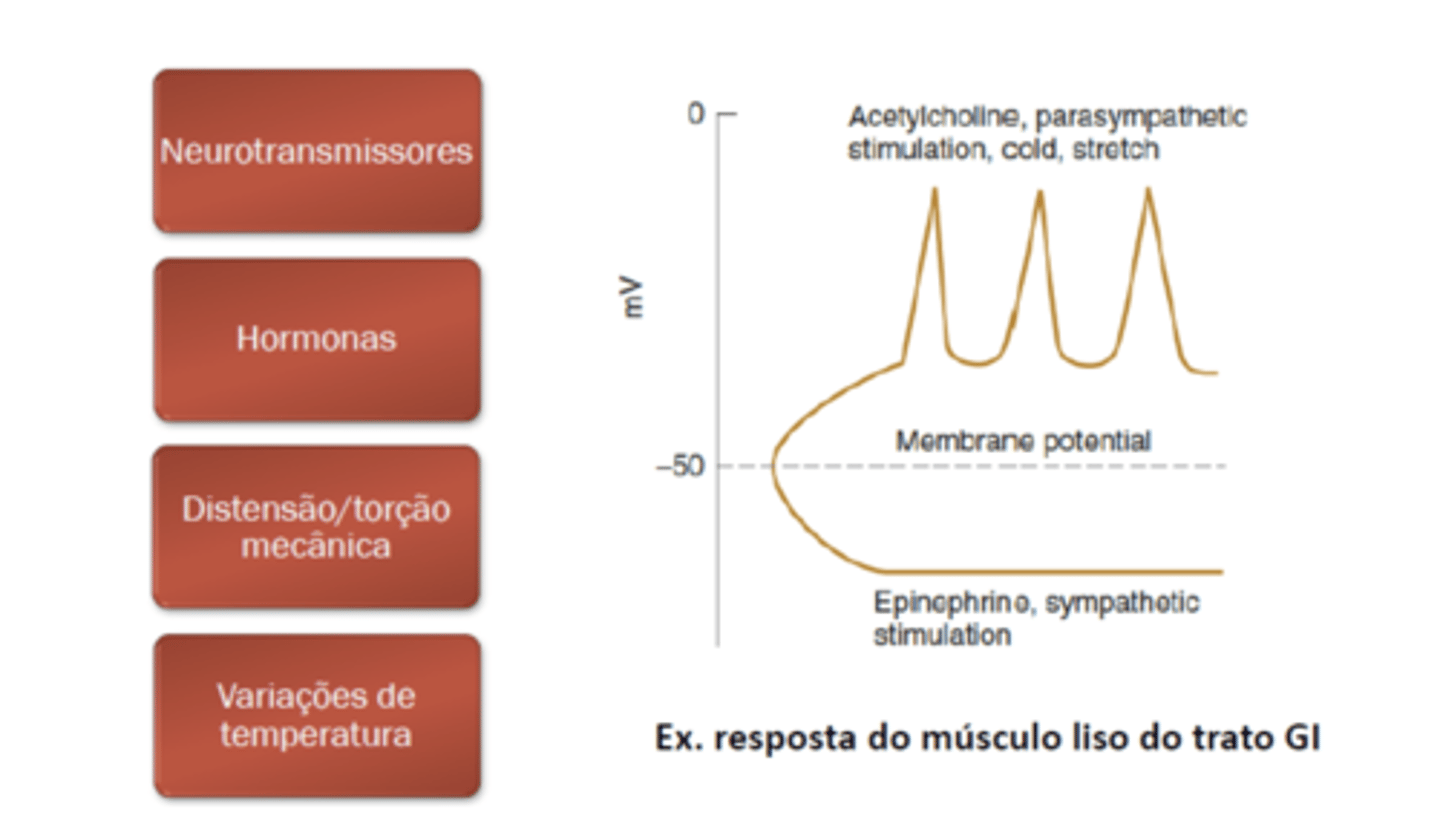
plateau
sao canais de clacio mt tempo abertos
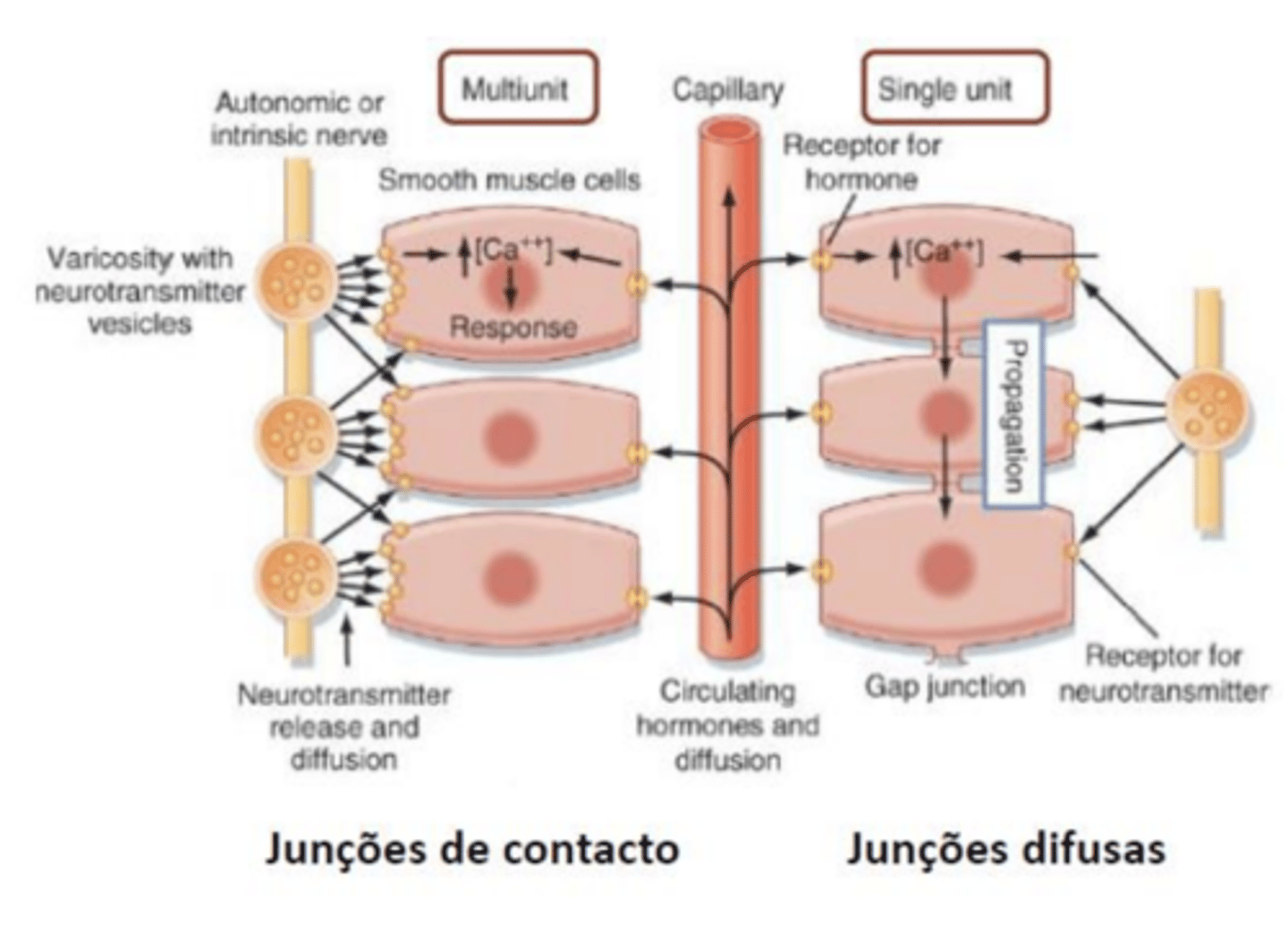
junções difusas
partilha citoplasma uma estimulação estimula todas as células
ou seja, o mesmo potencial de ação excita todas as células
junções de contacto
cada ramificação produz varicozidas onde se forma sinapses quimica de passagem
ocorre sinapse até ser produzida outra
há arias formas do clacio se armazenar de 2 formas diferentes, extra ou intracelularmente
cada células é excitada individualmente
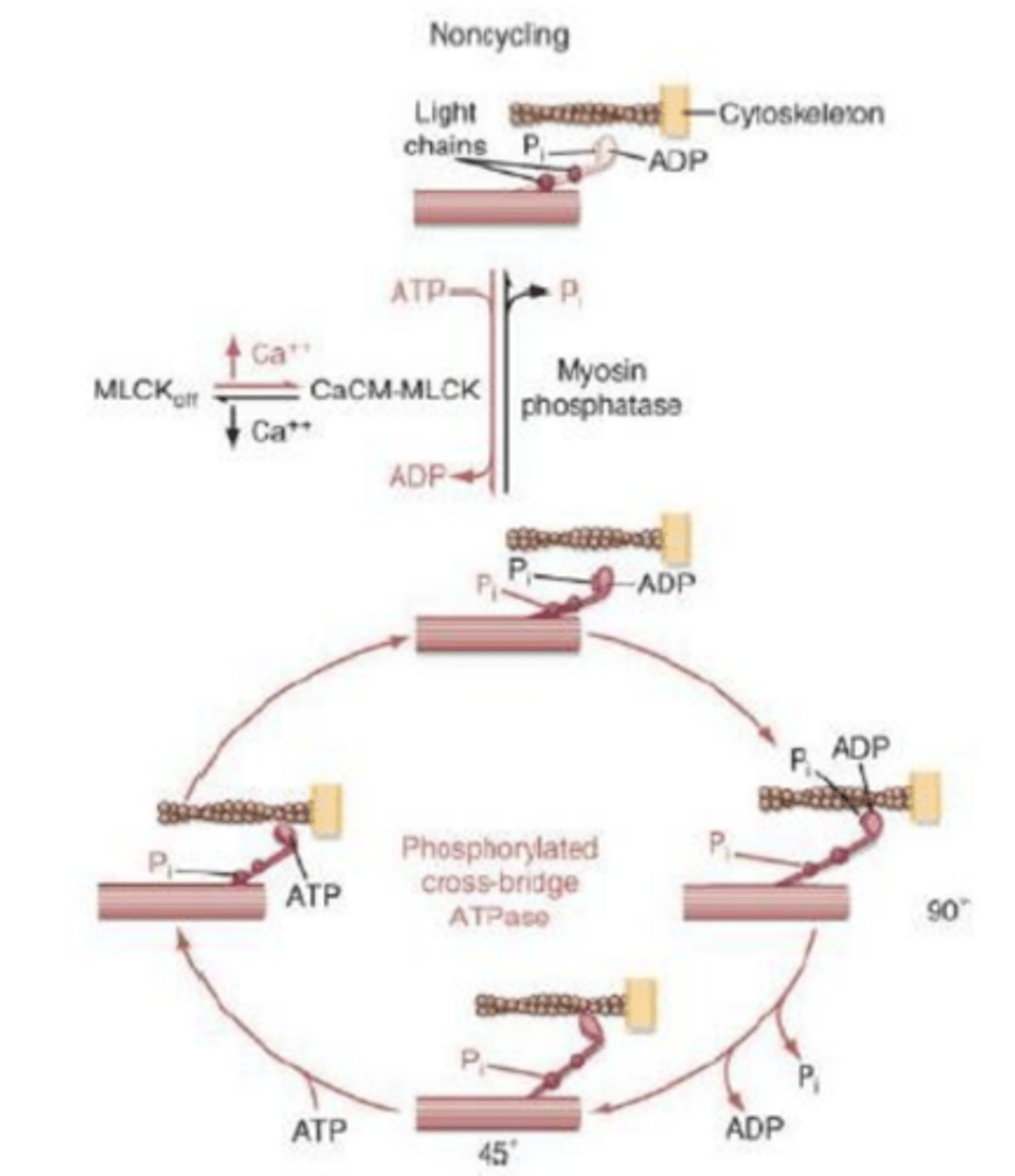
Relaxamento/Contração do Músculo Liso
O ciclo de contração-relaxamento é lento e é regulado pela miosina ATPase.
A remoção do cálcio é feita por bombas lentas e, portanto, o processo é todo bastante mais lento do que nos outros tipos de contração. Além disto, é um processo que usa bastante menos ATP (10 a 300 vezes menos) para manter o mesmo tipo de tensão.
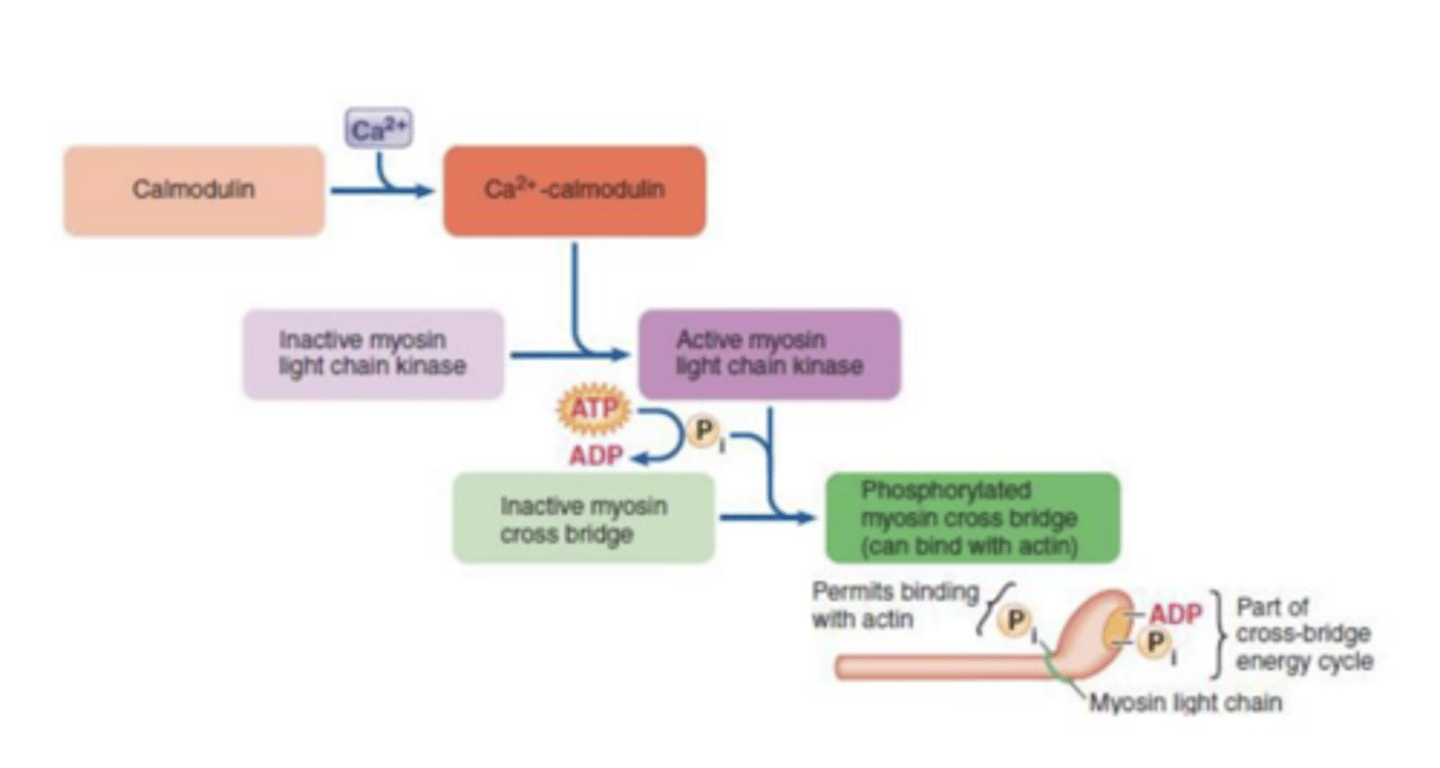
calmodulina q modula calcio subsitiui troponina
Excitação-contração mus liso
fosforilação da miosina possibilita ligação à actina através da calmodulia miosina tem q ser ativada
Relaxamento mus liso
fosfatase desfaz ligação da actina
fibras oxidativas lentas 1
treino de resistencia
maratonista
muita imoglubina
metabolismo oxidativo
cor vermelha
muita mioglobina
grande tamanho das unidades motoras
pouca velocidade de contração
atividade atpásica lenta
menor afinidade
demoram a fatigar
fibras oxidativas rápidas 2a
treino de não resistencia
fisioculturista
poucas mitocondria pq nao fazem fosforilação oxidativa
não são muito relevantes pois não existem em grande nº
fibras glicolíticas rápidas 2b
metabolismo glicolítico
cor branca
mioglobulina grande
contração rápida
maior ativiadade atpásica
fatigam rapidamente
Tipos de treino
hiperplasia -- podem causar hiperplasia muscular
atrofia
diminuição do volume do músculo por diminuição das fibras
Atrofia por desuso
• Via neuronal intacta
• Desuso prolongado (acamados,
membros imobilizados...)
Atrofia por desnervação
• Via neuronal está lesada
(traumatismo, infeção, neuropatia...)
Sistema propriocetivo
músculo tem sistmas de deteção das forças a q estão sujeitos
Fusos musculares - celulas sensoras, não contraem detatam variação de força
(fibras intrafusais)
- Distensão
- Comprimento
- Posição
- Inervados por fibras sensitivas Ia
Órgãos tendinosos de Golgi - deteta quantidade de força q músculo gera
- Força muscular
- Inervados por fibras sensitivas Ib e tb aalfa
Arco reflexo
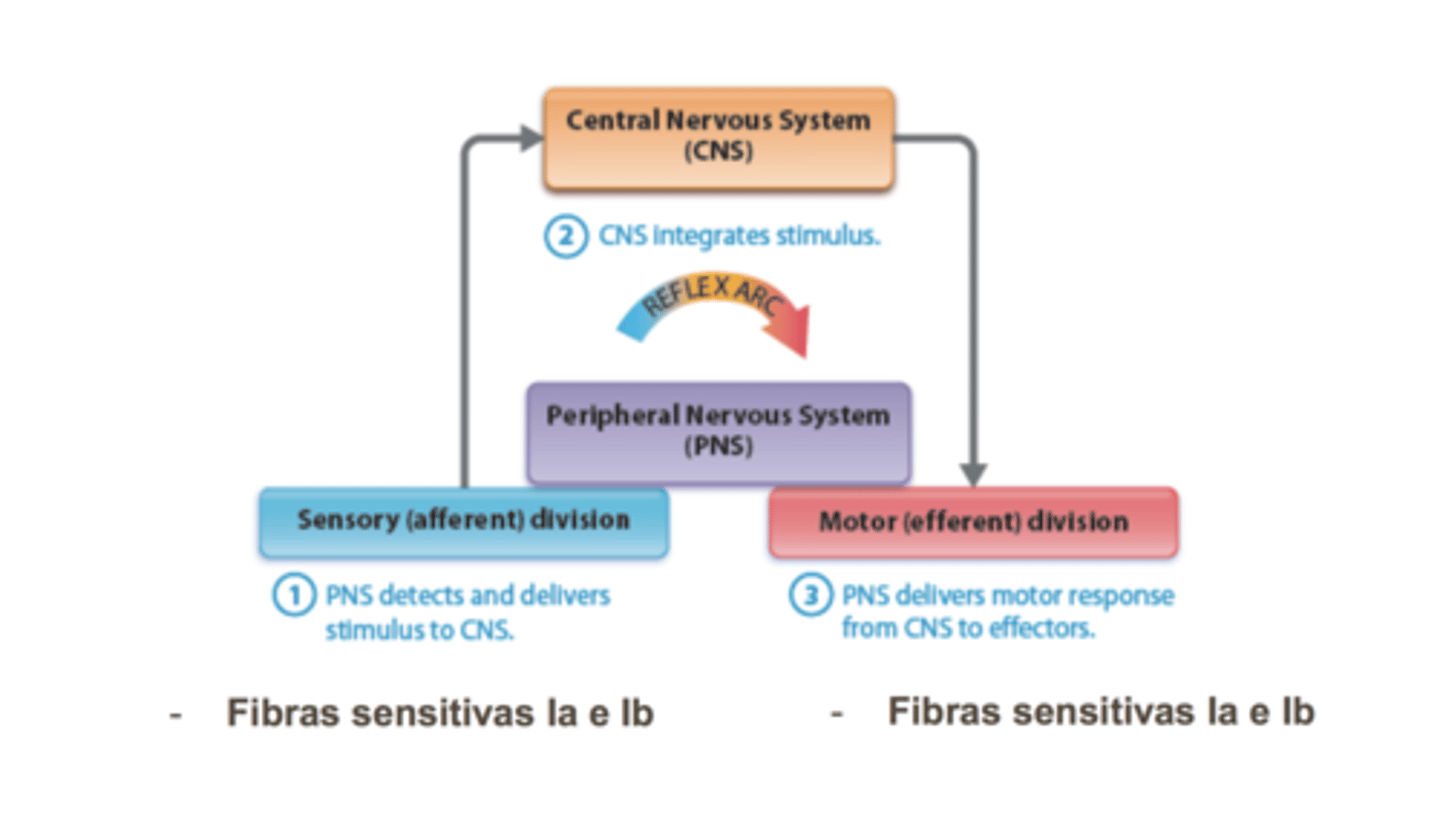
Reflexo miotático (de estiramento)
estiramento induz reposta de contração de encurtamente
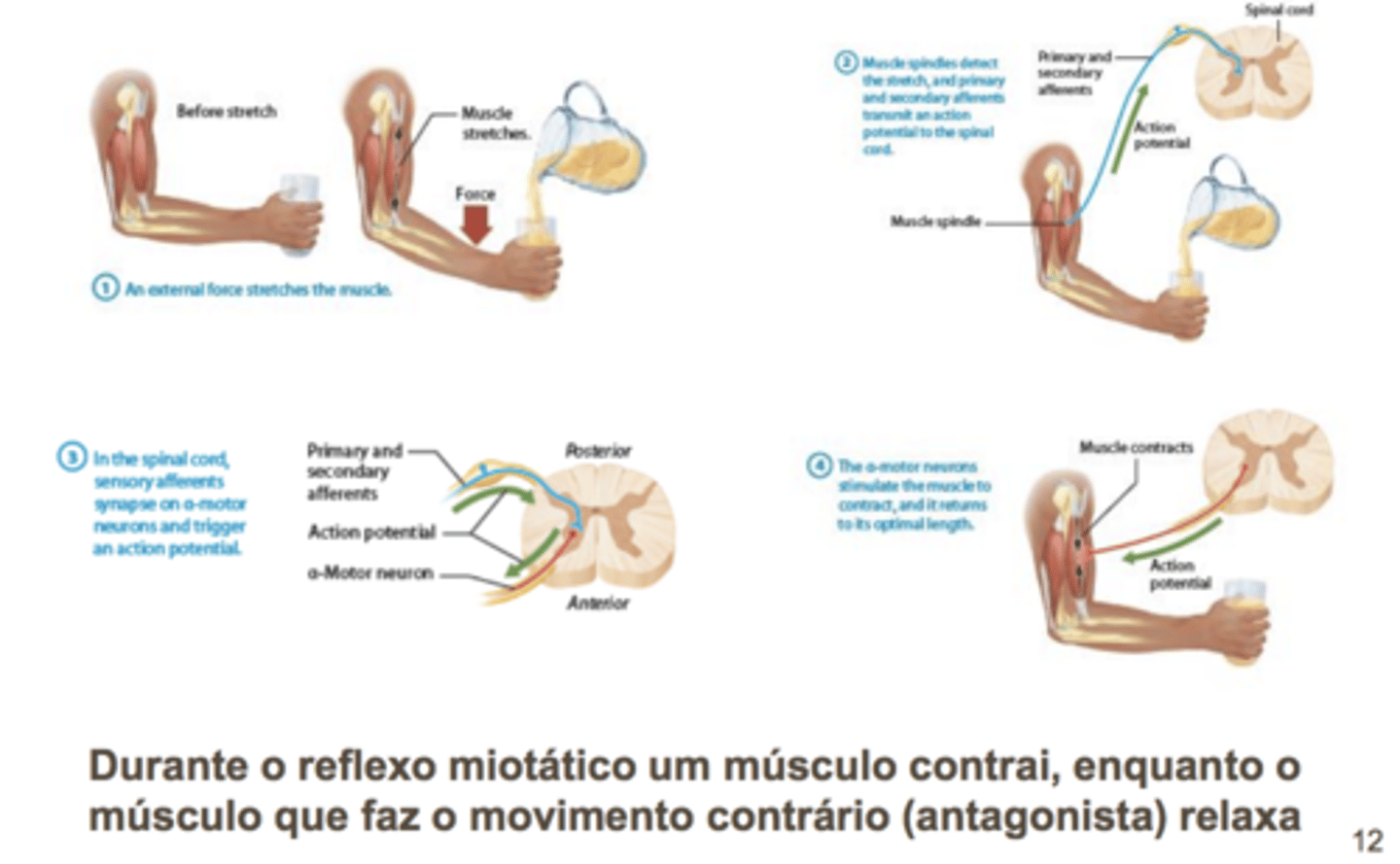
Reflexo miotático patelar
reflexo monocinático - força com encurtamento q causa extensão da perna, sinpase excitatoria com neurónio inibitorio na medula. Músculo no compartimento anterior da coxa sao encurtadores, sendo que no reflexo estes músculos sao inibidos
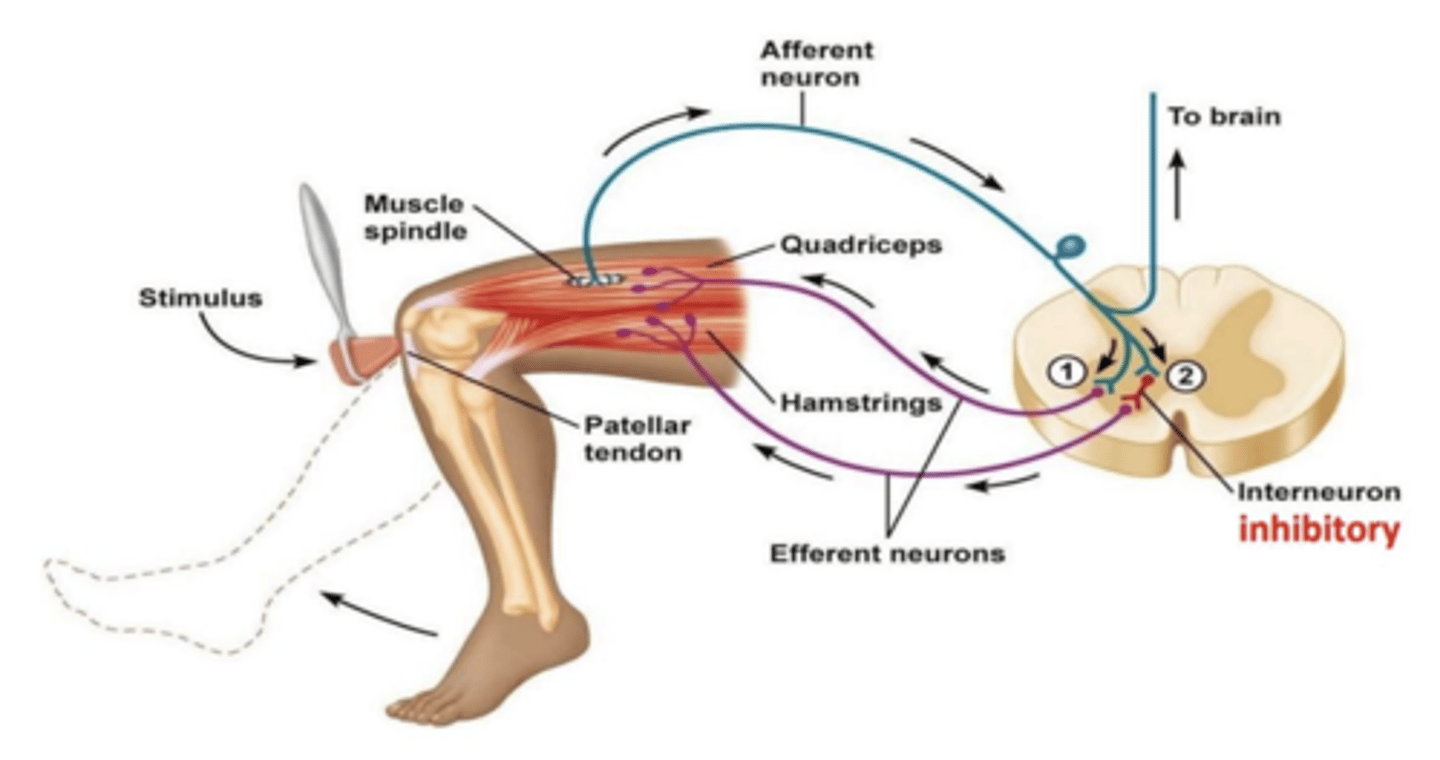
Reflexo miotático inverso (de Golgi)
quando músculo gera força excessiva, desse tendão surgem potencias de ação que vão à medula espinhais que fazem sinapse excitatória com interneurónio inibitório que induz inibição do músculo alfa motor travando o músculo - 1ª sinapse. 2ª sinapse com interneurónio excitatório do músculo antagonista
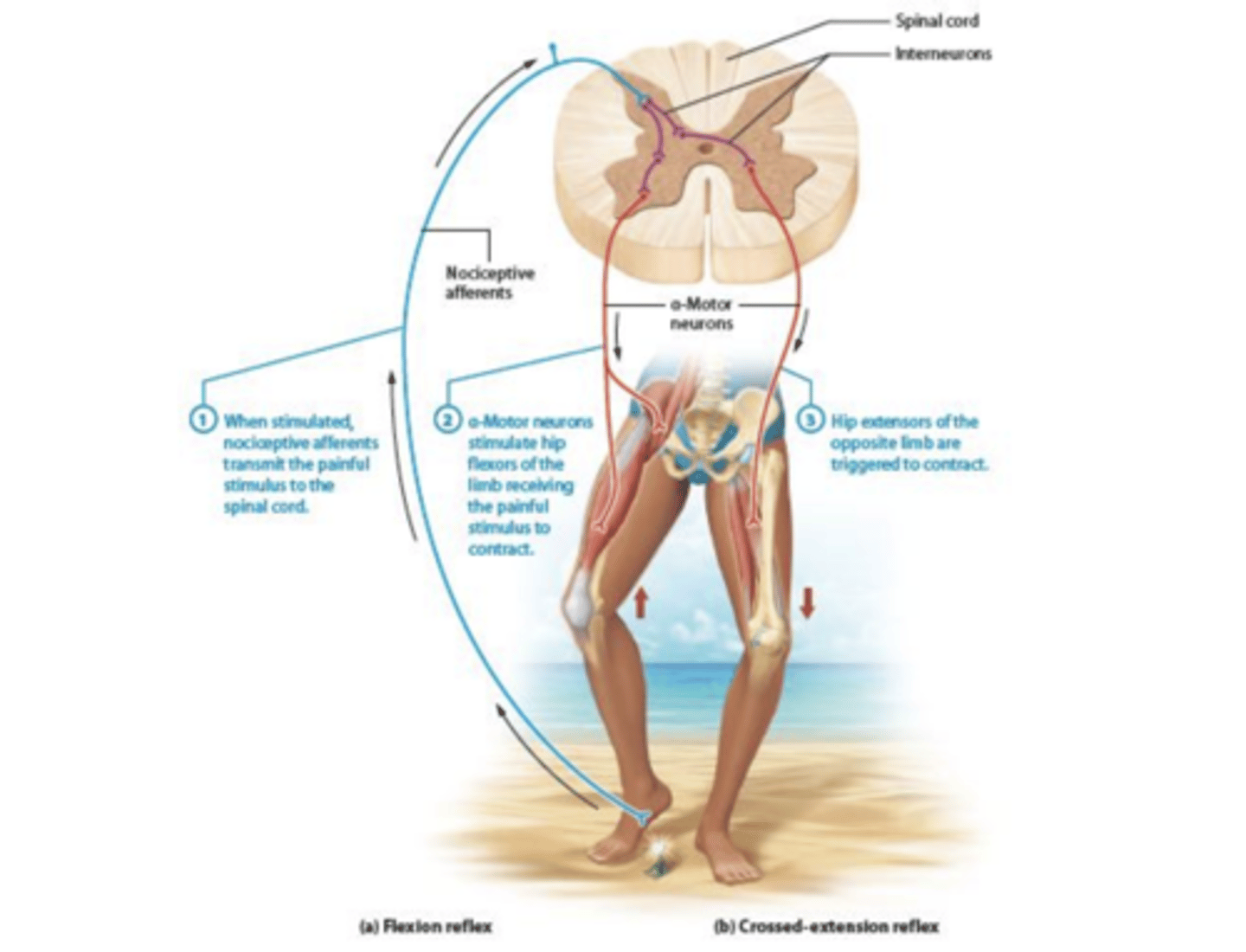
Reflexo de extensão cruzada
manter estabilidade dos membros inferior através do aumento da força do membro apoiado etimulaçãode 2 neuronios excitatórios
contração fásica
temporaria
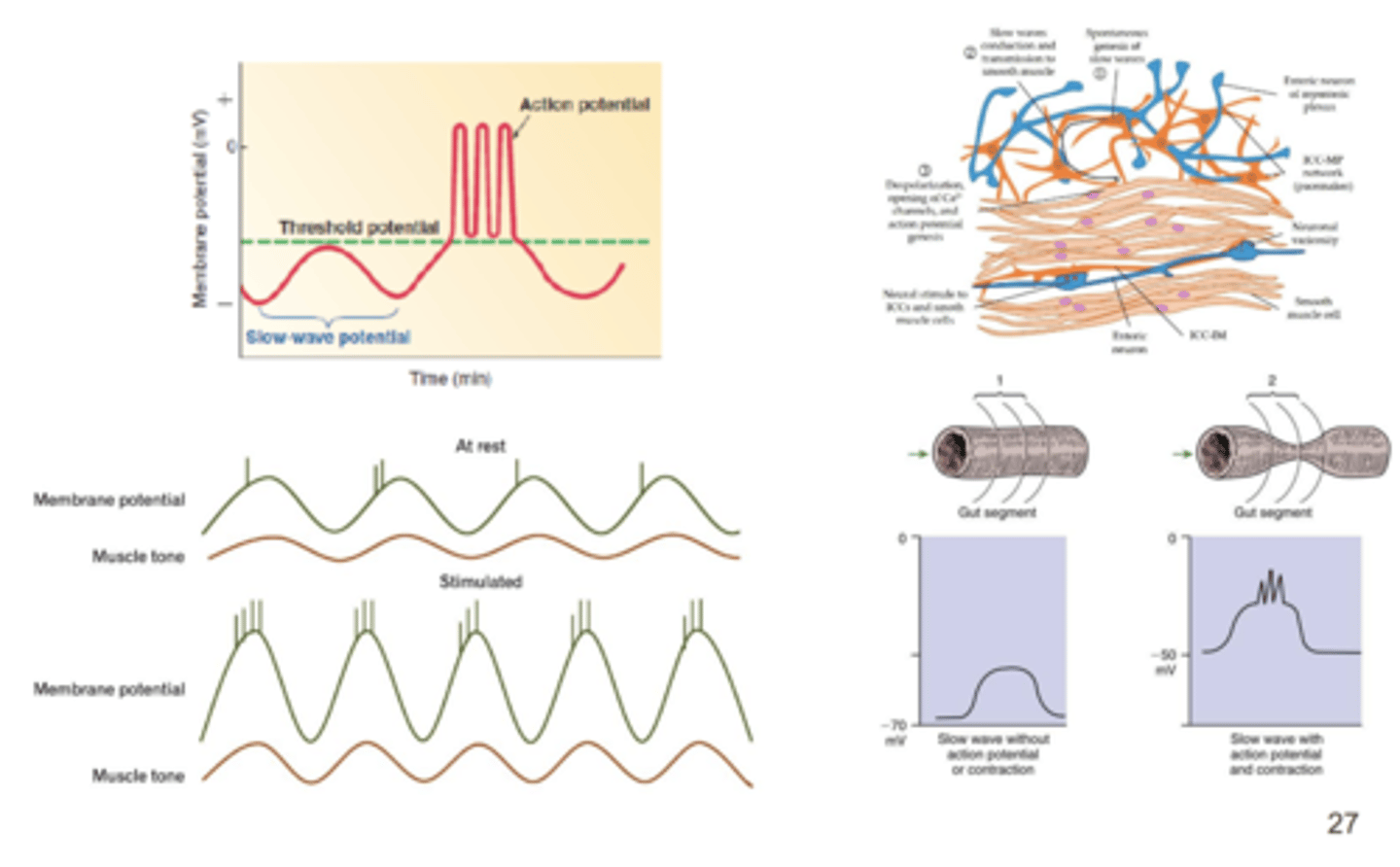
contração tónica
calibre constante
contração tipo plateau prolongada, há aumeo mantido calcio que entra q despolariza é usada para pontes cruzadas
contração tipo tónica
esfinter
contração tonica e distensão fásica
vasos
contração tónica
musc liso estomago e intestino
contração fásica, esporadiacamente contraem mesmo sem conteúdo para manutenção
bexiga, esófago
relaxada na maior parte do tempo, havendo contração fásica de vez em quando